КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Структура популяции 1 страница
|
|
|
|
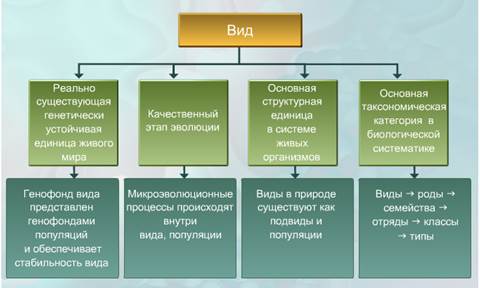
Понятие вида.Основной, элементарной и реально существующей единицей органического мира, или иначе — универсальной формой существования жизни, является вид (от лат. species — взгляд, образ).Вид — исторически сложившаяся совокупность популяций, особи которых обладают наследственным сходством морфологических, физиологических и биохимических особенностей, могут свободно скрещиваться и давать плодовитое потомство, приспособлены к определенным условиям жизни и занимают определенную область— ареал.
Особи, принадлежащие к одному виду, не скрещиваются с особями другого вида, характеризуются генетической общностью, единством происхождения. Вид существует во времени: он возникает, распространяется (в период расцвета), может сохраняться неопределенно долгое время в устойчивом, почти неизменном состоянии (реликтовые виды) или непрерывно изменяться. Одни виды со временем исчезают, не оставляя новых ветвей. Другие дают начало новым видам.
Вид как таксономическая категория. Вид — основная категория биологической классификации. Понятие «вид» впервые было введено в конце
17 в. английским ботаником Джоном Реем (1627—1709), отметившим, что разные виды отличаются по внешнему и внутреннему строению и не скрещиваются между собой.
Большой вклад в дальнейшую разработку понятия «вид» внес шведский ученый Карл Линней (1707—1778). Согласно его представлениям, виды — объективно существующие в природе образования, и между разными видами в большей или меньшей степени имеются различия (рис. 1.1). Так, например, явно различаются между собой по внешним признакам медведь и волк, в то время как волк, шакал, гиена, лисица внешне более сходны, так как принадлежат к одному семейству — волчьих. Еще в большей степени сходна внешность у видов одного рода. Вот почему вид начали рассматривать как основную классификационную единицу. Это имело огромное значение для развития систематики.
Таким образом, с именем Линнея связано начало описания и классификации живых организмов. Эта работа продолжается и в настоящее время.
Критерии вида. Признаки, по которым один вид можно отличить от другого, называют критериями вида.
В основе морфологического критерия лежит сходство внешнего и внутреннего строения между особями одного вида. Этот критерий — самый удобный и поэтому широко используется в систематике.
Однако особи в пределах вида иногда так сильно различаются, что только по морфологическому критерию не всегда удается определить, к какому виду они относятся. Вместе с тем существуют виды морфологически сходные, но особи этих видов не скрещиваются между собой. Это — виды-двойники, которые исследователи открывают во многих систематических группах. Так, под названием «крыса черная» различают два вида-двойника, имеющих в кариотипах по 38 и 42 хромосомы. Установлено также, что под названием «малярийный комар» существует до 15 внешне неразличимых видов, ранее считавшихся одним видом. Около 5% всех видов насекомых, птиц, рыб, земноводных, червей составляют виды-двойники.
В основу физиологического критерия положено сходство всех процессов жизнедеятельности у особей одного вида, прежде всего сходство размножения. Особи разных видов, как правило, не скрещиваются, или потомство их бесплодно. Например, у многих видов мухи дрозофилы сперма особей чужого вида вызывает иммунную реакцию, что приводит к гибели сперматозоидов в половых путях самки. В то же время в природе есть виды, особи которых скрещиваются и дают плодовитое потомство (некоторые виды канареек, зябликов, тополей, ив).
Географический критерий основан на том, что каждый вид занимает определенную территорию или акваторию, называемую ареалом. Он может быть большим или меньшим, прерывистым или сплошным (рис. 1.2). Однако огромное число видов имеет накладывающиеся или перекрывающиеся ареалы. Кроме того, существуют виды, не имеющие четких границ распространения, а также виды-космополиты, обитающие на огромных пространствах суши всех континентов или океана (например, растения — пастушья сумка, одуванчик лекарственный, виды рдестов, ряски, тростника, животные-синантропы — постельный клоп, рыжий таракан, комнатная муха). Поэтому географический критерий, как и другие, не является абсолютным.
Экологический критерий основан на том, что каждый вид может существовать только в определенных условиях, выполняя свойственные ему
функции в определенном биогеоценозе. Так, например, лютик едкий произрастает на пойменных лугах, лютик ползучий — по берегам рек и канав, лютик жгучий — на заболоченных местах. Существуют, однако, виды, которые не имеют строгой экологической приуроченности. К ним относятся многие сорные растения, а также виды, находящиеся под опекой человека: комнатные и культурные растения, домашние животные.
Генетический (цитоморфологический) критерий основан на различии видов по кариотипам, т.е. числу, форме и размерам хромосом. Для подавляющего большинства видов характерен строго определенный кариотип. Однако и этот критерий не является универсальным. Во-первых, у многих видов число хромосом одинаково и форма их сходна. Например, некоторые виды семейства бобовых имеют 22 хромосомы (2п = 22). Во-вторых, в пределах одного и того же вида могут встречаться особи с разным числом хромосом, что является результатом геномных мутаций (поли- или анеу-плоидия). Например, ива козья может иметь диплоидное (38) или тетра-плоидное (76) число хромосом.
Биохимический критерий позволяет различать виды по составу и структуре определенных белков, нуклеиновых кислот и др. Особи одного вида имеют сходную структуру ДНК, что обусловливает синтез одинаковых белков, отличающихся от белков другого вида. Вместе с тем у некоторых бактерий, грибов, высших растений состав ДНК оказался очень близким. Следовательно, есть виды-двойники и по биохимическим признакам.
Таким образом, только учет всех или большинства критериев позволяет отличить особей одного вида от другого.
Основной формой существования жизни и единицей классификации живых организмов является вид. Для выделения вида используется совокупность критериев: морфологический, физиологический, географический, экологический, генетический, биохимический. Вид является результатом длительной эволюции органического мира. Будучи генетически закрытой системой, он, тем не менее, исторически развивается и изменяется.
6. Популяция и ее структура.
1. Понятие «популяция». Вплоть до настоящего времени нет четкого определения понятия «популяция», которое бы удовлетворяло потребностям различных биологических направлений. Причиной этого является то, что генетики в понятие «популяция» вкладывают свой конкретный смысл (как совокупность свободно скрещивающихся особей), систематики подходят с позиций морфофункционального единства, биогеографы – с позиций исторически обособленной группы организмов, характерной для конкретного ландшафта или группы ландшафтов, и т.д. С другой стороны, понятие «популяция» строится в зависимости от применяемого подхода к ее изучению. В-третьих, и что, пожалуй, основное, споры ведутся о том, что же является наименьшей структурной единицей вида, ибо популяции могут подразделяться на микропопуляции или объединяться в более крупные популяционные системы. Мы с вами подойдем к определению понятия «популяция» с экологической точки зрения. Однако будем помнить, современный эколог должен обязательно учитывать популяцию не просто как некое сообщество особей одного вида, но и как строго организованную в структурном и функциональном плане единицу вида как в пространстве, так и во времени.
Популяцией в экологии называют совокупность особей одного вида, находящихся во взаимодействии между собой и населяющих общую территорию. Члены популяции оказывают друг на друга не меньшее воздействие, чем физико-химические факторы среды или обитающие рядом организмы других видов. Поскольку контакты между особями одной популяции происходят чаще, чем между особями разных популяций, то именно внутривидовые отношения являются более напряженными, чем межвидовые.
В популяциях проявляются в той или иной степени все формы биотических отношений, но наиболее ярко выражены мутуалистические и конкурентные. Мутуализм – крайняя степень ассоциации между особями, при которой каждый извлекает выгоду из связи с другим организмом. Специфические внутривидовые связи – это отношения, направленные на воспроизводство: между особями разных полов и между родительскими и дочерними поколениями.
Популяция представляет собой форму внутривидовой организации или форму существования вида, обеспечивающую наиболее полное использование данной группой особей природных ресурсов территории, к которой популяция приурочена. Биологическое значение популяции состоит в наиболее полном и рациональном использовании энергетических ресурсов, позволяющем обеспечить оставление потомства. При половом размножении обмен генами превращает популяцию в целостную генетическую систему. Если размножение происходит вегетативным путем (побегами, почками и т.д.) или другими способами, то популяция представляет собой систему клонов, или чистых линий, совместно использующих среду.
В современной биологии популяция рассматривается как элементарная единица в процессе эволюции, способная реагировать на изменения среды перестройкой своего генофонда.
Говоря об экологических популяциях, необходимо отметить большое разнообразие их масштабов. У одного и того же вида в разной среде популяции могут сильно различаться. Эти различия обусловлены:
а) площадью ареала популяции – они могут занимать территорию, сравнимую по площади с материком (популяции песца, кряквы), и могут ограничиваться несколькими квадратными метрами (некоторые амфибии и моллюски);
б) количеством особей, образующих популяцию, – популяция может объединять миллионы особей (комары) или всего несколько десятков животных (крупные хищники);
в) количеством микропопуляций – одни популяции представлены множеством микропопуляций, приуроченных к разным биотопам, другие – едины в пространственном отношении.
Таким образом, популяции представляют собой весьма разнообразные видовые группировки, количество и особенности которых соответствуют пестроте и условиям местообитания, специфическим свойствам среды и биологии самих животных.
Под демографической структурой популяции понимаютпрежде всего ее половой и возрастной состав. Кроме того, принято говорить о пространственной структуре популяции - то есть об особенностях размещения особей популяции в пространстве.
Знание структуры популяции позволяет исследователю сделать выводы о ее благополучии или неблагополучии. Например, если в популяции отсутствуют генеративные (то есть способные дать потомство) особи и при этом много старовозрастных (сенильных) особей, то можно сделать неблагоприятный прогноз. У такой популяции может не быть будущего. Структуру популяции желательно изучать в динамике: зная ее изменение в течение нескольких лет, можно намного более уверенно говорить о тех или иных тенденциях.
Возрастная структура популяции. Этот тип структуры связан с соотношением особей различных возврастов в популяции. Особи одного возраста принято объединять в когорты, то есть возрастные группы.
Возрастная структура популяций растений описана очень подробно. В ней выделяют (по Т.А. Роботнову) следующие возрасты (возрастные группы организмов):
латентный период - состояние семени;
прегенеративный период (включает состояния проростка, ювенильного растения, имматурного растения и виргинильного растения);
генеративный период (обычно подразделяется на три подпериода - молодых, зрелых и старых генеративных особей);
постгенеративный период (включает состояния субсенильного растения, сенильного растения и фазу отмирания).
Разумеется, при этом возникает проблема соотношения календарного и биологического возраста. Принадлежность к определенному возрастному состоянию определяется по степени выраженности определенных морфологических (например, степень расчлененности сложного листа) и физиологических (например, способность дать потомство) признаков. Таким образом фиксируется, прежде всего, биологический возраст особи. Биологический возраст имеет для эколога большее значение, так как именно он определяет роль особи в популяционных процессах. В то же время, как правило, существует взаимосвязь между биологическим и календарным возрастом.
В популяциях животных также можно выделить различные возрастные стадии. Например, насекомые, развивающиеся с полным метаморфозом, проходят стадии яйца, личинки, куколки, имаго (взрослого насекомого). У других животных (развивающихся без метаморфоза) также можно выделить различные возрастные состояния, хотя границы между ними могут быть и не настолько четкими.
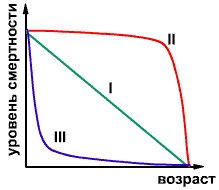
Характер возрастной структуры (или как говорят, возрастного спектра) популяции зависит от типа так называемой кривой выживания, свойственной данной популяции. Кривая выживания отражает уровень смертности в различных возрастных группах. Так, если уровень смертности не зависит от возраста особей, то кривая выживания представляет собой снижающуюся линию (см. рисунок, тип I). То есть отмирание особей происходит в данном типе равномерно, коэффициент смертности остается постоянным на протяжении всей жизни. Такая кривая выживания свойственна видам, развитие которых происходит без метаморфоза при достаточной устойчивости рождающегося потомства. Этот тип принято называть типом гидры - для нее свойственна кривая выживания, приближающаяся к прямой линии.
У видов, для которых роль внешних факторов в смертности невелика, кривая выживания характеризуется небольшим понижением до определенного возраста, после которого происходит резкое падение в следствие естественной (физиологический) смертности. Тип II на рисунке. Близкий к этому типу характер кривой выживания свойственен человеку (хотя кривая выживания человека несколько более пологая и, таким образом, является чем-то средним между типами I и II). Этот тип носит названия типа дрозофиллы: именно его демонстрируют дрозофиллы в лабораторных условиях (не поедаемые хищниками).
Для очень многих видов характерна высокая смертность на ранних стадиях онтогенеза. У таких видов кривая выживания характеризуется резким падением в области младших возрастов. Особи, пережившие "критический" возраст, демонстрируют низкую смертность и доживают до больших возрастов. Тип носит название типа устрицы. Тип III на рисунке.
Изучение кривых выживания представляет большой интерес для эколога. Оно позволяет судить о том, в каком возрасте тот или иной вид наиболее уязвим. Если действие причин, способных изменить рождаемость или смертность, приходится на наиболее уязвимую стадию, то их влияние на последующее развитие популяции будет наибольшим. Эту закономерность необходимо учитывать при организации охоты или в борьбе с вредителями.
Половая структура популяции. О половой структуре популяции можно говорить, разумеется, только если речь идет о раздельнополом (бисексуальном) виде. Бисексуальность играет огромную роль в поддержании генетической разнокачественности особей популяции. Значение генетической разнокачественности для устойчивости популяции будет подробно раскрыто в следующем уроке. Сейчас же отметим, что половая структура, то есть соотношение полов, имеет прямое отношение к воспроизводству популяции и ее устойчивости.
Принято выделять первичное, вторичное и третичное соотношение полов в популяции. Первичное соотношение полов определяется генетическими механизмами - равномерностью расхождения половых хромосом. Например, у человека XY-хромосомы определяют развитие мужского пола, а XX - женского. В этом случае первичное соотношение полов 1:1, то есть равновероятно.
Вторичное соотношение полов - это соотношение полов на момент рождения (среди новорожденных). Оно может существенно отличаться от первичного по целому ряду причин: избирательность яйцеклеток к сперматозоидам, несущим X- или Y-хромосому, неодинаковой способностью таких сперматозоидов к оплодотворению, различными внешними факторами. Например, зоологами описано влияние температуры на вторичное соотношение полов у рептилий. Аналогичная закономерность характерна и для некоторых насекомых. Так, у муравьев оплодотворение обеспечивается при температуре выше 20 С, а при более низких температурах откладываются неоплодотворенные яйца. Из последних вылупляются самцы, а из оплодотворенных - преимущественно, самки.
Третичное соотношение полов - это соотношение полов среди взрослых животных.
Пространственная структура популяции. Пространственная структура популяции отражает характер размещение особей в пространстве.
Выделяют три основных типа распределения особей в пространстве:
единообразное (особи размещены в пространстве равномерно, на одинаковых расстояниях друг от друга), тип также носит название равномерного распределения;
конгрегационное, или мозаичное (то есть "пятнистое", особи размещаются в обособленных скоплениях);
случайное, или диффузное (особи распределены в пространстве случайным образом).
Если вы дружны со статистикой, то различие между этими типами пространственной структуры можно описать так. Возьмем некоторое число выборок, подсчитав число особей на равных площадях. Если дисперсия числа особей в выборках стремится к нулю - мы имеем дело с равномерным распределением. Если дисперсия близка к среднему арифметическому - это случайное распределение. Если же дисперсия на много больше среднего арифметического, то можно говорить о конгреационном размещении особей.
Равномерное распределение встречается в природе редко и чаще всего вызвано острой внутривидовой конкуренцией (как, например, у хищных рыб).
Случайное распределение можно наблюдать только в однородной среде и только у видов, которые не обнаруживают никакого стремление к объединению в группы. Как хрестоматийный пример равномерного распределения, обычно приводят распределение жука Tribolium в муке.
Распределение группами встречается намного чаще. Оно связано с особенностями микросреды или с особенностями поведения животных.
Пространственная структура имеет важное экологическое значение. Прежде всего, определенный тип использования территории позволяет популяции эффективно использовать ресурсы среды и снизить внутривидовую конкуренцию. Эффективность использования среды и снижение конкуренции между представителями популяции позволяют ей укрепить свои позиции по отношению к другим видам, населяющим данную экосистему.
Другое важное значение пространственной структуры популяции состоит в том, что она обеспечивает взаимодействие особей внутри популяции. Без определенного уровня внутрипопуляционных контактов популяция не сможет выполнять как свои видовые функции (размножение, расселение), так и функции, связанные с участием в экосистеме (участие в круговоротах веществ, создание биологической продукции и так далее).
7. Характерные свойства популяций. Динамика плотности популяций.
Популяции состоят из множества особей. Особенности их состава изучает демография (буквально — «народоописание»), наука, методы которой развивались в первую очередь при описании динамики численности и состава поселений человека. Демографические характеристики популяций можно разделить на две группы: статические и динамические. Статические характеристики популяций могут быть определены для конкретного момента времени; примером такой характеристики может быть численность популяции. Как бы быстро ни изменялась эта величина, в любой момент мы могли бы установить, сколько особей входит в состав той или иной популяции. К этой категории, кроме численности популяций, относится ее состав (соотношение в ней особей разного возраста и пола), плотность (отношение численности популяции к занимаемой ею площади или объему), характерное распределение особей в пространстве и некоторые другие параметры.
Численность популяции может определяться различными способами. Для определения поголовья крупных, хорошо заметных организмов, образующих скопления на относительно небольших территориях, используют прямой подсчет. Так можно подсчитать колониально гнездящихся птиц (грачей, уток), копытных (северных оленей). В некоторых случаях эффективен способ мечения. При этом животных метят и выпускают туда же, где они были пойманы. Через некоторое время производят новый отлов в том же месте, и по доле меченых особей от общего числа отловленных особей определяют численность популяции. Так определяют численность земноводных, мелких птиц, мышевидных грызунов и многих других животных.
Но зачастую невозможно определить общую численность организмов прямым подсчетом особей. При этом приходится довольствоваться отбором проб и подсчетом количества особей в них. При этом измеряется плотность — число особей, приходящихся на единицу пространства. Плотность наземных организмов выражают на единицу площади, а популяций планктонных организмов — на единицу объема водной толщи. Иногда плотность оценивают как число встреч на маршруте. Так оценивают птиц по пению в весенний период, млекопитающих по следам в зимний период.
Еще одной статической характеристикой является пространственное распределение особей и их групп. Выделяют три основных типа пространственных распределений: случайное, регулярное и групповое. При случайном распределении нахождение каждой особи никак не зависит от расположения других особей. В природе оно встречается довольно редко.
Регулярное распределение наблюдается в том случае, когда между особями действуют силы отталкивания (например, для них характерна защита индивидуальной территории). В густом ельнике стволы отдельных деревьев удалены друг от друга на расстояние максимум двух крон, а одиноко стоящее дерево затеняет пространство вокруг ствола, предотвращая рост проростков и, соответственно, появление новых особей этого вида. Такое размещение часто встречается и в искусственно созданных экосистемах (парках, агросистемах).
При групповом распределении (пятнистом, агрегированном) вероятность нахождения пустых участков и участков с несколькими особями намного выше, чем при случайном распределении. Ярким его примером является расположение травянистых растений на болоте, когда они занимают возвышенные участки, образуя «пятнистые» скопления. Как и расположение отдельных особей, распределение групп может также быть случайным, регулярным или пятнистым.
Приведенным выше перечнем статические характеристики популяций не исчерпываются. Например, видам, для которых характерна внутрипопуляционная иерархия (порядок подчинения), важна иерархическая структура популяций — отражение статуса отдельных особей и их отношений друг с другом. А какие процессы приводят к изменению статических (т.е. вовсе не неизменяемых!) характеристик популяции? Рождаемость, смертность, миграции — динамические характеристики. Природа этих параметров такова, что они могут быть измерены только для определенного промежутка времени. Важнейшие динамические характеристики популяции могут быть объединены в следующую простую формулу:
Изменение численности = (рождаемость + иммиграция) — (смертность + эмиграция)
При рассмотрении изменения численности необходимо также учитывать продолжительность жизни особей рассматриваемой популяции. Для организмов каждого вида существует некая максимальная продолжительность жизни особей. Она чаще всего необходима как крайняя точка при построении кривых выживания и демографических таблиц. Так у бактерий она может составлять десятки минут, а у древесных растений — десятки столетий.
Как мы сказали, важнейшими статическими характеристиками популяции являются ее половой состав (соотношение особей разного пола) и возрастной состав (соотношение особей разного возраста). Эти параметры принято описывать с помощью демографических таблиц. Первая такая таблица была построена основателем демографии Джоном Грантом в XVII веке на основании данных о смертности жителей Лондона, которые церковные приходы собирали, чтобы вовремя обнаруживать начало эпидемий чумы.
Демографические таблицы удобны для наблюдения за динамикой рождаемости и смертности в разных возрастных и (или) половых группах. Одним из способов их построения (табл. 4.3.1) является наблюдение за судьбой определенной группы особей (когорты), рожденных за короткий промежуток времени, и регистрация возраста наступления смерти всех членов когорты.
Но чаще в демографии пользуются иным методом: определением смертности в разных возрастных группах в течение какого-то периода наблюдений (табл. 4.3.2). Зная численность отдельных групп, можно рассчитать смертность для каждого возраста. Этот метод позволяет оценить смертность и выживаемость у видов с большой продолжительностью жизни, даже используя только статистические данные, относящиеся к небольшому промежутку времени.
На основании демографических таблиц строят демографические пирамиды. На вертикальной шкале откладывают возрастные интервалы, по левой стороне, в виде столбчатой диаграммы, — численность самцов, по правой — самок. При этом наглядным становится разница смертности в разных возрастных категориях и у разных полов.
Демографические пирамиды помогают зримо представить историю популяции. Рассмотрите такую пирамиду для населения России (рис. 4.3.1). Вы можете увидеть, например, как на ней отражается снижение рождаемости во время Великой Отечественной войны. «Эхо» этих событий проявилось даже через поколение и, в меньшей степени, через два. Количество людей, которые по своему возрасту являются детьми и внуками тех, кто появлялся на свет во время войны, оказывается меньше численности близких к ним возрастных групп.
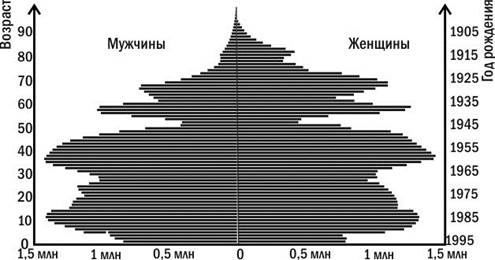
Рис. 4.3.1. Демографическая пирамида населения России состоянием на 1996 год
Демографические таблицы дают материал для построения кривых выживания. Такой способ графического представления зависимости доли сохранившихся в живых особей от их возраста в 20-х годах прошлого века предложил Роберт Перль. Он выделил три основных типа кривых выживания (рис. 4.3.2).
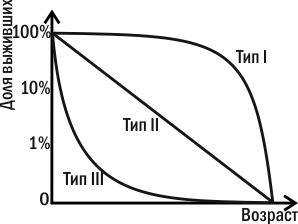
Рис. 4.3.2. Три типа «идеальных» кривых смертности по Перлу
Кривая I типа (типа дрозофилы) имеет выпуклую форму. Она описывает ситуацию, когда высокая смертность наблюдается в зрелом возрасте. Это характерно для дрозофил, поденок и других насекомых, которые вскоре после выхода из куколки оставляют потомство и погибают. К кривой I типа приближаются кривые выживания крупных млекопитающих.
Кривая II типа (типа гидры) характерна для организмов с равномерной смертностью в любом возрасте. На графике этому соответствует прямая линия. Такие типы кривых характерны для рыб, пресмыкающихся, птиц, травянистых многолетних растений и т.д., с единственной оговоркой, что отсчет ведется от организмов, уже прошедших наиболее уязвимые стадии своего развития.
Кривая III типа (типа устрицы) имеет вогнутую форму. Она характерна для организмов, которые в основном гибнут на начальных этапах своей жизни. Устрицы ведут прикрепленный образ жизни во взрослом состоянии, а личинка у них планктонная. Именно в этот период они наиболее уязвимы. У особей, удачно прошедших личиночную стадию, шанс выжить сильно повышается. Такой тип кривых выживания характерен для многих животных с высокой плодовитостью и отсутствием заботы о потомстве.
Реальные кривые выживания являются сочетанием этих типов. Кривая человека выпукла, относительно приближаясь к первому типу, однако в разных случаях может иметь разную форму (рис. 4.3.3).
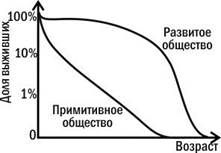
Рис. 4.3.3. Типы кривых выживания в примитивном и развитом обществе
В 1536 г. испанский аделантадо Педро де Мендоза, закладывая город Буэнос-Айрес, привез в аргентинские пампы 20 коров и 72 лошади. Спустя три года поселение было сожжено дотла индейцами, и испанцы его покинули. Лошади и коровы оказались предоставлены сами себе. Они размножились в пампах, и к 1700 г. численность популяции коров и популяции лошадей достигли миллиона голов каждая. Испанские мореплаватели XVI и XVII вв. систематически завозили на океанические острова коз, чтобы обеспечить себе пропитание на случай кораблекрушения. Один такой путешественник, Хуан Фернандес, завез пару коз на острова Тихого океана вблизи побережья Чили, — острова, которые затем были названы его именем. В 1704 г., когда Александр Селкирк (послуживший Даниэлю Дефо прототипом Робинзона Крузо) был оставлен на этих островах капитаном его корабля, численность стада коз, которым дала начало эта пара, превышала 10 000, и стадо существует до сих пор (О. Солбриг, Д. Солбриг, 1982).
Прирост популяции пропорционален ее численности, и поэтому, если рост популяции не ограничивают никакие внешние факторы, популяция растет ускоренно. Опишем этот рост математически.
Прирост популяции пропорционален численности особей в ней, то есть Δ N~N, где N — численность популяции, а Δ N — ее изменение за определенный период времени. Если этот период бесконечно мал, можно написать, что dN /dt=r×N, где dN/dt — изменение численности популяции (прирост), а r — репродуктивный потенциал, переменная, характеризующая способность популяции увеличивать свою численность. Приведенное уравнение называется экспоненциальной моделью роста численности популяции (рис. 4.4.1).
|
|
|
|
|
Дата добавления: 2015-04-24; Просмотров: 2734; Нарушение авторских прав?; Мы поможем в написании вашей работы!