КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Структура популяции 2 страница
|
|
|
|

Рис.4.4.1. Экспоненциальный рост
Величину r называют иногда мальтузианским параметром. Английский священник Томас Мальтус был первым, кто обратил внимание на то, что численность населения растет в геометрической прогрессии. Именно знакомство с его работой подтолкнуло и Чарльза Дарвина, и Альфреда Уоллеса к догадке о том, что потомство любых организмов должно «прореживаться» естественным отбором.
Как легко понять, с ростом времени численность популяции растет все быстрее, и достаточно скоро устремляется к бесконечности. Естественно, никакое местообитание не выдержит существования популяции с бесконечной численностью. Тем не менее, существует целый ряд процессов популяционного роста, который в определенном временном промежутке может быть описан с помощью экспоненциальной модели. Речь идет о случаях нелимитированного роста, когда какая-то популяция заселяет среду с избытком свободного ресурса: коровы и лошади заселяют пампу, мучные хрущаки — элеватор с зерном, дрожжи — бутыль виноградного сока и т.д.
Естественно, экспоненциальный рост популяции не может быть вечным. Рано или поздно ресурс исчерпается, и рост популяции затормозится. Каким будет это торможение? Практическая экология знает самые разные варианты: и резкий взлет численности с последующим вымиранием популяции, исчерпавшей свои ресурсы, и постепенное торможение прироста по мере приближения к определенному уровню. Проще всего описать медленное торможение. Простейшая описывающая такую динамику модель называется логистической и предложена (для описания роста численности популяции человека) французским математиком Ферхюльстом еще в 1845 году. В 1925 году аналогичная закономерность была заново открыта американским экологом Р. Перлем, который предположил, что она носит всеобщий характер.
В логистической модели вводится переменная K — емкость среды, равновесная численность популяции, при которой она потребляет все имеющиеся ресурсы. Прирост в логистической модели описывается уравнением dN /dt=r×N × (K-N)/K (рис. 4.4.2).

Рис. 4.4.2. Логистический рост
Пока N невелико, на прирост популяции основное влияние оказывает сомножитель rN и рост популяции ускоряется. Когда становится достаточно высоким, на численность популяции начинает оказывать основное влияние сомножитель (K-N)/K и рост популяции начинает замедляться. Когда N=K, (K-N)/K=0 и рост численности популяции прекращается.
При всей своей простоте логистическое уравнение удовлетворительно описывает много наблюдаемых в природе случаев и до сих пор с успехом используется в математической экологии.
8. Экологическая ниша.
Для понимания различного вида существующих связей в экосистемах и обусловленности механизмов их функционирования важно познакомиться с одним из основополагающих понятий экологии - экологической нишей.
Каждый вид или его части (популяции, группировки различного ранга) занимают определенное место в окружающей их среде. Например, определенньш вид животного не может произвольно менять пищевой рацион или время питания, место размножения, убежища и т. п. Для растений подобная обусловленность условий выражается, например, через светолюбие или тенелюбие, место в вертикальном расчленении сообщества (приуроченность к определенному ярусу), время наиболее активной вегетации. Например, под пологом леса одни растения успевают закончить основной жизненный цикл, завершающийся созреванием семян, до распускания листьев древесного полога (весенние эфемеры). В более позднее время их место занимают другие, более теневыносливые растения. Особая группа растений способна на быстрый захват свободного пространства (растения-пионеры), но отличается низкой конкурентной способностью и поэтому быстро уступает свое место другим (более конкурентоспособным) видам.
Приведенные примеры иллюстрируют экологическую нишу или отдельные ее элементы. Под экологической нишей понимают обычно место организма в природе и весь образ его жизнедеятельности, или, как говорят, жизненный статус, включающий отношение к факторам среды, видам пищи, времени и способам питания, местам размножения, укрытий и т. п. Это понятие значительно объемнее и содержательнее понятия «местообитание». Американский эколог Одум образно назвал местообитание «адресом» организма (вида), а экологическую нишу - его «профессией». На одном местообитании живет, как правило, большое количество организмов разных видов. Например, смешанный лес - это местообитание для сотен видов растений и животных, но у каждого из них своя и только одна «профессия» - экологическая ниша. Так, сходное местообитание, как отмечалось выше, в лесу занимают лось и белка. Но ниши их совершенно разные: белка живет в основном в кронах деревьев, питается семенами и плодами, там же размножается и т. п. Весь жизненный цикл лося связан с подпологовым пространством: питание зелеными растениями или их частями, размножение и укрытие в зарослях и т. п.
Если организмы занимают разные экологические ниши, они не вступают обычно в конкурентные отношения, сферы их деятельности и влияния разделены. В таком случае отношения рассматриваются как нейтральные.
Вместе с тем в каждой экосистеме имеются виды, которые претендуют на одну и ту же нишу или ее элементы (пищу, укрытия и пр.). В таком случае неизбежна конкуренция, борьба за обладание нишей. Эволюционно взаимоотношения сложились так, что виды со сходными требованиями к среде не могут длительно существовать совместно. Эта закономерность не без исключений, но она настолько объективна, что сформулирована в виде положения, которое получило название «правило конкурентного исключения». Автор этого правила эколог Г. Ф. Гаузе. Звучит оно так: если два вида со сходными требованиями к среде (питанию, поведению, местам размножения и т. п.) вступают в конкурентные отношения, то один из них должен погибнуть либо изменить свой образ жизни и занять новую экологическую нишу. Иногда, например, чтобы снять острые конкурентные отношения, одному организму (животному) достаточно изменить время питания, не меняя самого вида пищи (если конкуренция возникает на почке пищевых отношений), или найти новое местообитание (если конкуренция имеет место на почве данного фактора) и т. п.
Из других свойств экологических ниш отметим, что организм (вид) может их менять на протяжении своего жизненного цикла. Наиболее яркий пример в этом отношении - насекомые. Так, экологическая ниша личинок майского жука связана с почвой, питанием корневыми системами растений. В то же время экологическая ниша жуков связана с наземной средой, питанием зелеными частями растений.
Сообщества (биоценозы, экосистемы) формируются по принципу заполнения экологических ниш. В природном сформировавшемся сообществе обычно все ниши заняты. Именно в такие сообщества, например в долгосуществующие (коренные) леса, вероятность внедрения новых видов очень мала. В то же время следует иметь в виду, что занятость экологических ниш в определенной мере понятие относительное. Все ниши обычно освоены теми организмами, которые характерны для данного региона. Но если организм приходит извне (например, заносятся семена или другие зачатки) случайно или преднамеренно, например в результате внедрения человеком новых видов (интродукция, акклиматизация), то он может найти для себя свободную нишу в связи с тем, что на нее не было претендентов из набора существующих видов. В таком случае обычно неизбежно быстрое увеличение численности (вспышка) вида-пришельца, поскольку он находит крайне благоприятные условия (свободную нишу) и, в частности, не имеет врагов (хищников, паразитов или других организмов, которые им питаются). Такие явления не единичны. Например, размножение кроликов, завезенных в Австралию; перемещение ондатры из Азии в европейскую часть; интенсивное продвижение колорадского жука в новые районы.
С экологическими нишами в значительной мере связаны жизненные формы организмов. К последним относят группы видов, часто систематически далеко отстоящие, но выработавшие одинаковые морфологические адаптации в результате существования в сходных условиях. Например, сходством жизненных форм характеризуются дельфины (млекопитающие) и интенсивно передвигающиеся в водной среде хищные рыбы. В условиях степей сходными жизненными формами представлены тушканчики и кенгуру (прыгуны). В растительном мире отдельными жизненными формами представлены многочисленные виды деревьев, занимающие в качестве нити верхний ярус, кустарники, существующие под пологом леса, и травы - в напочвенном покрове.
9. Структура биогеоценоза. Ярусность. Экосистемы.
Сообщества организмов тесно связаны не только друг с другом, но и с абиотической средой. Растения могут существовать только при наличии света, углекислого газа, воды, минеральных солей. Животные и другие гетеротрофные организмы (грибы, большинство бактерий) живут за счет автотрофов, но нуждаются в поступлении таких неорганических соединений, как кислород и вода. В любом биотопе запасы неорганических соединений, необходимых для поддержания жизнедеятельности населяющих его организмов, сравнительно малы и постоянно убывают, поэтому необходимо их возобновление. Из окружающей среды живые организмы поглощают биогенные элементы и энергию и возвращают их обратно (например, при дыхании, выделении экскрементов, разложении растительных и животных остатков). Благодаря этим обменным процессам биоценоз и окружающая его неорганическая среда (экотоп) представляют собой сложную систему, получившую название экосистема или биогеоценоз.
Термин «экосистема» был предложен в 1935 г. английским экологом А. Тенсли, который подчеркивал, что в природе органические (биотические) и неорганические (абиотические) факторы выступают как равноправные компоненты и не следует отделять организмы от окружающей их среды.
Таким образом, биогеоценоз — это однородный участок земной поверхности с определенным составом живых организмов (биоценоз) и определенными условиями среды обитания (биотоп), которые объединены обменом веществ и энергии в единый природный комплекс (рис. 14.3). Во многих странах мира такие природные комплексы называют экологическими системами (экосистемами).
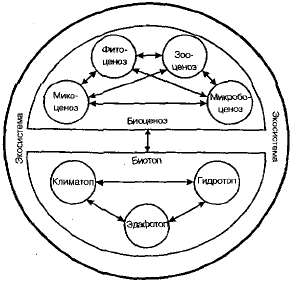
Рис. 14.3. Биоценоз в экосистеме.
Биогеоценоз и экосистема — понятия сходные, но не тождественные. Понятие «экосистема» не имеет ранга и размерности, поэтому оно применимо как к простым (муравейник, гниющий пень) и искусственным (аквариум, водохранилище, парк), так и к сложным естественным комплексам организмов с их средой обитания. Биогеоценоз, согласно российскому ученому В. Н, Сукачеву, отличается от экосистемы определенностью объема. Если экосистема может охватывать пространство любой протяженности. -— от капли прудовой воды с содержащимися в ней микроорганизмами до биосферы в целом, то биогеоценоз —- это экосистема, границы которой обусловлены характером растительного покрова, т. е. определенным фитоценозом. Следовательно, любой биогеоценоз является экосистемой, но не всякая экосистема есть биогеоценоз.
В основе концепции экосистемы лежит взаимосвязь между физическими и биологическими явлениями, между живой и косной материями. Хотя представление о структурных единицах природы формировалось в среде философов и естествоиспытателей веками, термин экосистема ввел в 1935г. английский ботаник А. Тэнсли (A.G. Tansley), понимавший под ним единый комплекс живых существ и абиотических условий, приуроченный к территории, занятой конкретным фитоценозом.
Согласно Тенсли, в экосистему входит «не только комплекс организмов, но и весь комплекс физических факторов, образующих то, что мы называем средой обитания, - факторы местообитания в самом широком смысле. Хотя главным интересующим нас объектом могут быть организмы, однако, когда мы пытаемся проникнуть в самую суть вещей, мы не можем отделить организмы от их особой среды, в сочетании с которой они образуют некую физическую систему» (цит. по [5]).
Следует отметить, что благодаря избирательной способности организмов к условиям среды заселение территории происходит в строгом соответствии с этими условиями и каждому местообитанию, как правило, соответствует один биоценоз.
Экосистема - основная функциональная единица в экологии, и для понимания проблем, возникающих в биосфере, необходимо прежде всего изучать экосистемный уровень организации. Поскольку экосистема представляет собой открытую систему, важной составляющей этого изучения являются процессы на входе и выходе из нее.
Наряду с термином «экосистема» широкое распространение получил другой термин - «биогеоценоз», введенный русским ученым, лесным экологом академиком В.Н. Сукачевым (1944), основоположником биогеоценологии. Биогеоценоз представляет собой участок территории (или акватории), через который не проходит ни одна установимая существенная биоценотическая, почвенно-геохимическая, геоморфологическая (раздельно учитывая границы векторов поверхностного и грунтового стоков) и микроклиматическая границы. Он представляет собой далее неподразделимую единицу биогеохимической работы [10].
Вертикальные границы биогеоценоза возможно установить следующие. За нижнюю границу как для практических, так и для теоретических целей достаточно принять нижнюю границу верхнего водоносного слоя (не верховодки, а постоянного уровня грунтовой воды); верхней границей следует считать нижние слои тропосферы, находящиеся в постоянном газообмене с почвой, растительным покровом и животным населением. Предварительная наметка границ биогеоценоза, необходимая для дальнейшего комплексного их определения и изучения, может быть проведена в согласии с высказыванием В.Н. Сукачева, путем установления природных фитоценологических границ.
В современной литературе термин «экосистема» стал пониматься очень широко - как любое сочетание организмов со средой, и, следовательно, экосистема может быть единицей и меньшей, чем биогеоценоз (типа кочки на болоте, поляны в лесу, соответствующей понятию «парцелла», по Н.В. Дылису), и гораздо большей, вплоть до биосферы в целом.
Структура экосистемы - сложное многоплановое понятие. Ее определяют как пространственно-временную организацию (упорядоченность) объекта. Понятие структуры имеет три аспекта, соответствующие трем этапам развития и усложнения этого понятия. Первоначальное представление сводилось к тому, что под структурой понималось взаимное расположение составных частей. В этом представлении заключен лишь чисто пространственный аспект структуры. При дальнейшем развитии понятия возник его функциональный аспект, который требует обращать внимание на способы соединения частей системы, то есть на внутренние системообразующие связи. Однако представление о структуре экосистемы оставалось статичным, пока не появился третий, динамический, или временной, аспект, то есть структура экосистемы стала рассматриваться не только как некоторая организованность его составных частей в пространстве, но и как упорядоченность смены его состояний во времени [2].
Важно подчеркнуть, что экосистемный уровень рассмотрения подразумевает наличие лишь вертикальной структуры. Горизонтальная (пространственная) структура рассматривается на других уровнях иерархии природных систем (ландшафты, природные территориальные комплексы, биосфера в целом).
Структурными элементами экосистемы являются абиотические (косные) компоненты среды (воздух, воды, литогенная основа), биокосные (почва) и живые организмы (растения, животные, грибы, микроорганизмы). Пространственная упорядоченность структуры выражается в закономерном расположении компонентов по вертикали и существовании между ними вертикальных же потоков вещества и энергии.
С точки зрения трофической1 структуры экосистему можно разделить на два яруса: верхний автотрофный (самостоятельно питающийся) ярус,включающий растения1, в которых благодаря фотосинтезу используются простые неорганические вещества и происходит накопление сложных богатых энергией органических соединений; нижний гетеротрофный (питаемый другими) ярус, в котором преобладают использование, трансформация и разложение сложных, накопленных в автотрофном ярусе соединений.
Разнообразие экосистем отражает количество разных типов местообитаний, сообществ и экологических процессов и является одним из уровней биоразнообразия2. Под биологическим разнообразием понимают число различимых типов биологических объектов или явлений и частоту их встречаемости в фиксированном интервале пространства и времени, в общем случае отражающих сложность живого вещества, способность его к саморегуляции своих функций и возможность его разностороннего использования. Биоразнообразие включает в себя все виды животных, растений грибов и микроорганизмов, экосистем и протекающие в них процессы.
В экосистеме можно выделить два компонента — биотический и абиотический. Биотический делится на автотрофный и гетеротрофный компоненты, формирующие трофическую структуру экосистемы.
Единственным источником энергии для существования экосистемы и поддержания в ней различных процессов являются продуценты, усваивающее энергию солнца, с эффективностью 0,1 — 1 %, редко 3 — 4,5 % от первоначального количества. Автотрофы представляют первый трофический уровень экосистемы. Последующие трофические уровни экосистемы формируются за счёт консументов и замыкаются редуцентами, которые переводят неживое органическое вещество в минеральную форму, которая может быть усвоена автотрофным элементом.
Основные компоненты экосистемы
С точки зрения структуры в экосистеме выделяют:
климатический режим, определяющий температуру, влажность, режим освещения и прочие физические характеристики среды;
неорганические вещества, включающиеся в круговорот;
органические соединения, которые связывают биотическую и абиотическую части в круговороте вещества и энергии;
продуценты — организмы, создающие первичную продукцию;
макроконсументы, или фаготрофы, — гетеротрофы, поедающие другие организмы или крупные частицы органического вещества;
микроконсументы — гетеротрофы, в основном грибы и бактерии, которые разрушают мёртвое органическое вещество, минерализуя его, тем самым возвращая в круговорот.
Последние три компонента формируют биомассу экосистемы.
С точки зрения функционирования экосистемы выделяют следующие функциональные блоки организмов:
биофаги — организмы, поедающие других живых организмов,
сапрофаги — организмы, поедающие мёртвое органическое вещество.
Данное разделение показывает временно-функциональную связь в экосистеме, фокусируясь на разделении во времени образования органического вещества и перераспределении его внутри экосистемы и переработки сапрофагами. Между отмиранием органического вещества и повторным включением его составляющих в круговорот вещества в экосистеме может пройти существенный промежуток времени, например, в случае соснового бревна, 100 и более лет.
Все эти компоненты взаимосвязаны в пространстве и времени и образуют единую структурно-функциональную систему.
Эта структура сообщества определяется, прежде всего, сложением фитоценоза. Как правило, фитоценозы расчленены на достаточно хорошо отграниченные в пространстве (по вертикали и по горизонтали), а иногда и во времени элементы структуры. К таким элементам относятся ярусы и микрогруппировки. Первые характеризуют вертикальное, вторые — горизонтальное расчленение фитоценозов.
Основной фактор, определяющий вертикальное распределение растений, — количество света, обусловливающее температурный режим и режим влажности на разных уровнях над поверхностью почвы. Растения верхних ярусов более светолюбивы, чем низкорослые, и лучше них приспособлены к колебаниям температуры и влажности воздуха; нижние ярусы образованы растениями менее требовательными к свету; травянистый покров леса в результате отмирания листьев, стеблей, корней участвует в процессе почвообразования и тем самым влияет на растения верхнего яруса.
Ярусы (I—V) особенно хорошо заметны в лесах умеренного пояса (рис. 2). В них можно выделить 5-6 ярусов: первый (верхний) ярус образуют деревья первой величины (дуб черешчатый, липа сердцевидная, вяз гладкий и др.); второй — деревья второй величины, а также кустарник (рябина обыкновенная, дикие яблоня и груша, черемуха и др.); третий ярус состоит из высоких трав (чистец лесной, крапива, сныть обыкновенная) и кустарничков (черника); четвертый ярус состоит из высоких трав (чистец лесной, крапива, сныть обыкновенная) и кустарничков (черника); пятый ярус сложен из низких трав (осока волосистая, копытень европейский); в шестом ярусе — мхи, лишайники.
Животные также преимущественно приурочены к тому или иному ярусу растительности. Например, среди птиц есть виды, гнездящиеся только на земле (фазановые, тетеревиные, трясогузки, коньки, овсянки), другие — в кустарниковом ярусе (дрозды, славки, снегири) или в кронах деревьев (зяблики, щеглы, корольки, крупные хищники и др.).
Расчлененность (неоднородность) в горизонтальном направлении — мозаичность — свойственна практически всем сообществам. Мозаичность выражается наличием в биогеоценозе различных микрогруппировок, которые различаются видовым составом, количественным соотношением разных видов, сомкнутостью, продуктивностью и другими признаками и свойствами.
Неравномерность в распределении видов живых организмов в пределах сообществ и связанная с этим мозаичность обусловлены рядом причин: особенностями биологии размножения и формы растений, неоднородностью почвенных условий (наличие понижений и повышений), средообразующим влиянием растений и др. Мозаичность может возникнуть в результате деятельности животных (образованием муравейников, вытаптыванием травостоя копытными и др.) или человека (выборочная рубка, кострища и т. д.).
10. Основные характеристики экосистем: биомасса, биопродуктивность, энергетика, биоразнообразие.
Одно из важнейших свойств организмов, их популяций и экосистем в целом - способность создавать органическое вещество, которое называют продукцией. Образование продукции в единицу времени (час, сутки, год) на единице площади (метры квадратные, гектар) или объема (в водных экосистемах) характеризует продуктивность экосистем. Продукция и продуктивность могут определяться для экосистем в целом или для отдельных групп организмов (растений, животных, микроорганизмов) или видов.
Продукцию растений называют первичной, а животных - вторичной. Наряду с продукцией различают биомассу организма, групп организмов или экосистем в целом. Под ней понимают всю живую органическую массу, которая содержится в экосистеме или ее элементах вне зависимости от того, за какой период она образовалась и накопилась. Биомасса и продукция (продуктивность) обычно выражаются через абсолютно сухой вес.
Нетрудно понять, что величина биомассы экосистем или их звеньев во многом зависит не столько от их продуктивности, сколько от продолжительности жизни организмов и экосистем в целом. Например, большая биомасса характерна для лесных экосистем: в тропических лесах она достигает 800-1000 т/га, в лесах умеренной зоны -300-400 т/га, а в травянистых сообществах обычно не выходит за пределы 3-5 т/га. В то же время лесные и травянистые (например, луговые) экосистемы в сходных условиях существования по продуктивности могут мало различаться или различаются в сторону большей продуктивности как лесных, так и травянистых сообществ.
Для экосистем, представленных однолетними организмами, их годичная продуктивность и биомасса практически совпадают. Для древесных сообществ они резко различаются. Вообще соотношение биомассы и годовой продукции экосистем можно выразить формулой:
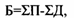
где Б - биомасса в данный момент времени, П - годовая продукция, Д - дыхание. Под последним применительно к экосистемам понимается вся сумма живого вещества, отчуждаемого на процессы разложения в результате гибели целых организмов (отпад) или их частей - сучьев, коры, листьев, наружных покровов (опад) и потребления гетеротрофами.
Энергия в экосистемах.
Напомним, что экосистема - это совокупность живых организмов, обменивающихся непрерывно энергией, веществом и информацией друг с другом и с окружающей средой. Рассмотрим сначала процесс обмена энергией. Энергию определяют как способность производить работу. Свойства энергии описываются законами термодинамики.
Первый закон (начало) термодинамики или закон сохранения энергии утверждает, что энергия может переходить из одной формы в другую, но она не исчезает и не создается заново. Второй закон (начало) термодинамики или закон энтропии утверждает, что в замкнутой системе энтропия может только возрастать. Применительно к энергии в экосистемах удобна следующая формулировка: процессы, связанные с превращениями энергии, могут происходить самопроизвольно только при условии, что энергия переходит из концентрированной формы в рассеянную, то есть деградирует. Мера количества энергии, которая становится недоступной для использования, или иначе мера изменения упорядоченности, которая происходит при деградации энергии, есть энтропия. Чем выше упорядоченность системы, тем меньше ее энтропия. Таким образом, любая живая система, в том числе и экосистема, поддерживает свою жизнедеятельность благодаря, во-первых, наличию в окружающей среде в избытке даровой энергии (энергия Солнца); во вторых, способности за счет устройства составляющих ее компонентов эту энергию улавливать и концентрировать, а использовав - рассеивать в окружающую среду. Таким образом, сначала улавливание, а затем концентрирование энергии с переходом от одного трофического уровня к другому обеспечивает повышение упорядоченности, организации живой системы, то есть уменьшение ее энтропии.
Энергия и продуктивность экосистем
Итак, жизнь в экосистеме поддерживается благодаря непрекращающемуся прохождению через живое вещество энергии, передаваемой от одного трофического уровня к другому; при этом происходит постоянное превращение энергии из одних форм в другие. Кроме того, при превращениях энергии часть ее теряется в виде тепла.
Тогда возникает вопрос: в каких количественных соотношениях, пропорциях должны находиться между собой члены сообщества разных трофических уровней в экосистеме, чтобы обеспечивать свою потребность в энергии?
Весь запас энергии сосредоточен в массе органического вещества - биомассе, поэтому интенсивность образования и разрушения органического вещества на каждом из уровней определяется прохождением энергии через экосистему (биомассу всегда можно выразить в единицах энергии). Скорость образования органического вещества называют продуктивностью. Различают первичную и вторичную продуктивность. В любой экосистеме происходит образование биомассы и ее разрушение, причем эти процессы всецело определяются жизнью низшего трофического уровня - продуцентами. Все остальные организмы только потребляют уже созданное растениями органическое вещество и, следовательно, общая продуктивность экосистемы от них не зависит. Высокие скорости продуцирования биомассы наблюдаются в естественных и искусственных экосистемах там, где благоприятны абиотические факторы, и особенно при поступлении дополнительной энергии извне, что уменьшает собственные затраты системы на поддержание жизнедеятельности. Такая дополнительная энергия может поступать в разной форме: например, на возделываемом поле - в форме энергии ископаемого топлива и работы, совершаемой человеком или животным. Таким образом, для обеспечения энергией всех особей сообщества живых организмов экосистемы необходимо определенноеколичественное соотношение между продуцентами, консументами разных порядков, детритофагами и редуцентами. Однако для жизнедеятельности любых организмов, а значит и системы в целом, только энергии недостаточно, они обязательно должны получать различные минеральные компоненты, микроэлементы, органические вещества, необходимые для построения молекул живого вещества.
В современных условиях роста антропогенной трансформации растительного покрова наиболее остро стоит проблема сохранениябиоразнообразия экосистемы как на видовом, так и на ценотическом уровнях. Познание закономерностей формирования структуры и динамики сообществ под влиянием природных и антропогенных факторов является узловым вопросом современной экологии и может служить теоретической базой для оценки состояния и прогноза изменений растительного покрова, а также планирования природоохранных мероприятий.
|
|
|
|
|
Дата добавления: 2015-04-24; Просмотров: 993; Нарушение авторских прав?; Мы поможем в написании вашей работы!