КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Зона поглощения веществ и дифференциации постоянных тканей
|
|
|
|
Зона роста
Зона роста представляет собой участок корня от зоны деления до корневых волосков. В этой зоне клетки сильно удлиняются путем растяжения. Переход от меристематической зоны к зоне роста постепенный. Интенсивность растяжения клеток на протяжении всей зоны роста имеет характер одновершинной кривой. Близ меристематической зоны она небольшая, затем резко увеличивается, достигая максимума, а потом снова постепенно уменьшается. Такие же особенности роста свойственны и междоузлиям стебля. Однако между стеблем и корнем есть и различия. В стебле апикальный рост сочетается с интеркалярным, в корне интеркалярного роста нет.
Эта зона обычно расположена на расстоянии 0,1-10 мм от кончика корня. Общая ее протяженность у разных растений варьирует от одного до нескольких сантиметров.
Первая постоянная, хотя и недолговечная ткань, дифференцирующаяся в корне, — ризодерма, или эпиблема, несущая корневые волоски. У некоторых папоротников ризодерма состоит из крупных бесцветных клеток с толстыми ослизняющимися оболочками, не образующих волоски, у ужовников развиваются очень редкие и короткие волоски. У большинства высших споровых и всех голосеменных растений ризодерма состоит из двух типов клеток: образующих волоски трихобластов и атрихобластов (см. гл. 5, п. 2).
Корневой волосок развивается очень быстро: у сеянцев яблони, например, — в течение 36-40 час. Длина волосков обычно составляет 0,15-8 мм, у травянистых растений они длиннее, чем у древесных. У сахарной свеклы волоски бывают длиной до 10 мм, у ясеня — 0,45-0,5 мм, у тепличных сеянцев цитрусовых они очень короткие — 0,05 мм. Волоски недолговечны. Через несколько дней они отмирают.
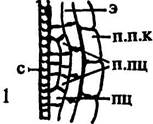
Число корневых волосков на 1 мм2 волосконосной зоны зависит от индивидуальных особенностей растений, а также от влажности почвы. У гороха их обычно 232, у проростков кукурузы —- 425, у сеянцев яблони — около 300. Волосков много у растений, живущих на достаточно увлажненных почвах, у водных растений они немногочисленные или отсутствуют. У кувшинки и кубышки абсорбирующая поверхность увеличивается не в результате развития волосков, а вследствие ветвления корней. Обычно волоски не развиваются на придаточных корнях некоторых луковичных растений, произрастающих на сильно увлажненном субстрате. Иногда их не бывает и у растений, развивающихся на сухих почвах, например, у шафрана, некоторых видов лютика. Волоски очень сильно увеличивают абсорбирующую поверхность корней. У сеянцев некоторых сортов яблони общее число волосков составляет 17,5 млн, их суммарная длина — 3 км, а у экземпляров пшеницы, культивируемых в благоприятных условиях, она нередко достигает 10000 км. Волоски активно поглощают воду с растворенными в ней минеральными веществами. В меньшей степени этой способностью обладают и ниже расположенные зоны корня, что связано с большой плотностью цитоплазмы их клеток. Ионы минеральных веществ, поглощенные меристемой и зоной роста, включаются в так называемый конструктивный обмен, то есть эти вещества используются для синтеза аминокислот, белков и других соединений, необходимых для увеличения общего количества цитоплазмы, а ионы, поступающие в корень через волоски, попадая в дифференцирующиеся сосуды, распространяются по всему растению, осуществляя, таким образом, минеральное питание. При этом поглощение почвенных растворов и их передвижение вглубь корня происходит и по апопласту, то есть по межфибриллярным пространствам оболочек и межклетникам, и по симпласту, то есть по протопластам клеток, используя цитоплазматические коммуникации, представленные плазмодесмами (рис. 163).

| Рис. 163. Строение корня в зоне корневых волосков. Обозначения: к.в — корневой волосок, мж — межклетники, мкс — метаксилема, п.к — первичная кора, пкс — протоксилема, пц — перицикл, рз — ризодерма, фл — флоэма, э — эндодерма с поясками Каспар и. Стрелками показаны пути передвижения растворов веществ, поглощаемых из почвы. Сплошными стрелками отмечены пути прохождения растворов по симпласту, пунктирными — по апопласту Не случайно поэтому дифференциация ксилемы начинается почти одновременно с развитием корневых волосков. В этой зоне начинается восходящий ток веществ. Как и в стебле, развитие флоэмы в корне опережает развитие ксилемы. Дифференцированные элементы этой ткани выявляются уже в зоне роста, в которой заканчивается нисходящий ток веществ, предста-вленных продуктами ассимиляции, используемыми для синтеза химических соединений, без которых невозможно увеличение количества цитоплазмы в растущих клетках. |
Развитию проводящих тканей предшествует обособление анатомо-то-пографических зон корня: первичной коры и центрального цилиндра, или стелы.
3.5. Зона первичного строения
Первичная кора состоит из паренхимных клеток, число слоев которых сильно варьирует. Наружный слой первичной коры после отмирания корневых волосков и сбрасывания ризодермы дифференцируется в первичную покровную ткань — экзодерму, а из внутреннего слоя развивается эндодерма.
Экзодерма (см. гл. 5, п. 1.2.), как особая ткань, клетки которой имеют опробковевшие оболочки, не выражена у споровых архегониальных растений. У семенных растений она выполняет функции защиты тканей корня от потери воды и растворенных в ней веществ, а также от проникновения в этот орган микроорганизмов. Ее строение варьирует у разных растений. У большинства двудольных, корни которых рано претерпевают вторичное утолщение, сопровождающееся отмиранием и сбрасыванием всей первичной коры, она обычно однослойная. В корнях однодольных экзодерма нередко многослойная (рис. 165 А) и содержит пропускные клетки, оболочки которых не опробковевают.
Эндодерма в зоне корневых волосков состоит из удлиненных тонкостенных клеток с поясками Каспари (см. гл. 7, п. 1), «опоясывающими» клетку посередине радиальных и поперечных стенок. На поперечных срезах корня пояски Каспари видны крайне редко, но обычно хорошо заметны сечения этих поясков на смежных радиальных стенках — пятна Каспари.
Так как субериноподобные вещества непроницаемы для воды, пояски Каспари препятствуют свободной диффузии ионов по оболочке, а плотное прилегание к ним плазмалеммы исключает возможность их передвижения в пространстве между нею и оболочкой. В связи с этим транспорт ионов всегда находится под контролем цитоплазмы самой клетки эндодермы.
Типичной эндодермы нет у плаунов, у представителей остальных систематических групп высших растений она может иметь разное строение.
Наличие поясков Каспари характерно для первой стадии развития эндодермы. Такую эндодерму имеют корни многих двудольных с рано начинающимся вторичным утолщением. У некоторых из них эндодерма достигает второй стадии развития, которая характеризуется отложением тонкого слоя субериноподобных веществ на внутренней поверхности всех клеточных стенок. Наконец, у однодольных растений, корни которых не имеют вторичного утолщения, эндодерма может переходить в третью стадию развития. В этом случае боковые и внутренние стенки клеток сильно утолщаются и одревесневают, их протопласты отмирают. У многих однодольных эндодерма дифференцирована на 2 типа клеток: мертвые, с U-образными утолщениями оболочек на поперечных срезах корня (см. рис. 165 В), играющие, по-видимому, механическую роль, и пропускные, осуществляющие физиологическую связь между первичной корой и центральным цилиндром. Обычно они расположены против элементов протоксилемы (рис. 165 Б). U-образные утолщения оболочек встречаются иногда и в клетках экзодермы, и в эндодерме других органов. Например, в корневищах ландыша они могут быть в клетках первичной коры, непосредственно прилегающих к эндодерме.
Средняя часть коры обычно многослойная, внутренние ее клетки у некоторых растений могут делиться периклинально, и их производные, размеры которых постепенно увеличиваются к периферии корня, растягиваясь при этом в тангентальном направлении, располагаются радиальными рядами. У растений из влажных местообитаний, а также у многих пальм в первичной коре часто образуются рексигенные воздухоносные полости. У пальм и некоторых других растений в ней встречаются группы волокон, некоторые клетки первичной коры склерифицируются (рис. 165 В).
3.6. Стела корня
Центральный цилиндр, или стела, состоит из перицикла и сложного радиального проводящего пучка, в котором радиальные тяжи первичной флоэмы чередуются по кругу с радиальными тяжами первичной ксилемы (рис. 34 Е; рис.165).
В корне развивается единственный тяж прокамбия. Первые элементы протофлоэмы закладываются экзархно, то есть в наружной части прокамбия. Между группами этих элементов так же экзархно закладываются элементы протоксилемы с кольчатыми и спиральными утолщениями оболочек. Дальнейшее развитие этих тканей осуществляется центростремительно: с внутренней стороны от элементов протофлоэмы и протоксилемы дифференцируются более широкопросветные элементы метафлоэмы и метаксилемы.
Тяжи ксилемы и флоэмы равномерно распределены по окружности центрального цилиндра. Число тяжей ксилемы всегда равно числу тяжей флоэмы, но у разных растений эти числа варьируют. В связи с этим различают корни монархине (от греч. arche — начало), имеющие по одному тяжу ксилемы и флоэмы, диархные, с двумя тяжами каждой из проводящих тканей, триархные, тетрархные и т. д. Если тяжей проводящих тканей много, проводящую систему называют полиархной.
Монархные корни встречаются крайне редко, они характерны для некоторых видов ужовника из отдела папоротниковидных. Наиболее распространены диархные корни. Полиархные корни свойственны, главным образом, однодольным.
От особенностей расположения элементов метаксилемы зависит общий вид центрального цилиндра на поперечном срезе корня.
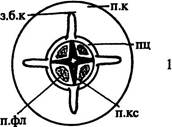
В корнях лука (рис. 165), тыквы и других растений наиболее широкопро-светный и самый молодой по времени образования водопроводящий элемент метаксилемы находится в центре (в зависимости от систематического положения растения он может быть представлен либо сосудом, либо трахеидой). На поперечном срезе такого корня ксилема имеет вид звезды с радиально расходящимися лучами, внутренние части которых состоят из метаксилемы, а наружные — из протоксилемы. Если корень диархный, то ксилема составляет пластинку, с каждой стороны от которой (но не по краям!) находится по одному тяжу флоэмы (см. рис. 192 А, 193 А, 194).
У некоторых растений формирование проводящих элементов происходит не по всей толщине прокамбия, а только в его наружной части. В этом случае внутренние клетки прокамбия дифференцируются в паренхимные клетки (кукуруза) или в волокна, оболочки которых впоследствии утолщаются и одревесневают (ирис). Эту внутреннюю часть центрального цилиндра нельзя называть по аналогии со стеблем сердцевиной, так как сердцевина стебля развивается из основной меристемы — производной медуллярной зоны конуса нарастания побега, а в корне она имеет прокамбиальное происхождение.
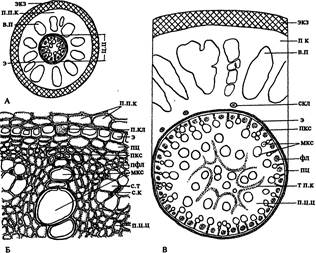
| Рис. 164. Строение тонкого (А) и толстого (В) корней пальмы хамеропс на поперечных срезах и фрагмент центрального цилиндра (Б). Обозначения: в.п — воздухоносные полости, мкс — метаксилема, п.к — первичная кора, п.кл — пропускная клетка, п.п.к — паренхимные клетки первичной коры, пфл — протофлоэма, п.ц.ц — паренхима центрального цилиндра, с.к — сопровождающая клетка, скл — склереиды, с.т — ситовидная трубка метафлоэмы, т.п.к — тяжи сплюснутых паренхимных клеток, фл — флоэма, ц.ц — центральный цилиндр, э — эндодерма, экз — экзодерма |
В корнях пальм (рис. 164 В) и некоторых других растений с полиархной проводящей системой во внутренней паренхиме или механической ткани центрального цилиндра могут быть и диффузно расположенные, местами контактирующие широкопросветные элементы метаксилемы. Некоторые из них примыкают к коротким радиальным тяжам ксилемы, находящимся в наружной части стелы.
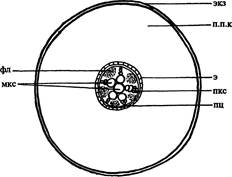
| Рис. 165. Строение корня лука репчатого на поперечном срезе (схема). Обозначения: мкс — метаксилема, пкс — протоксилема, п.п.к — паренхима первичной коры, пц — перицикл, фл — флоэма, экз — экзодерма, э — эндодерма |
Ксилема и флоэма состоят не только из проводящих элементов. В состав флоэмы входят паренхимные клетки. У бобовых, мальвовых и представителей других семейств цветковых в протофлоэме обычно развиваются волокна (см. рис. 167). В ксилеме могут быть элементы либриформа и мелкие паренхимные клетки, которые с возрастом одревесневают.
Перицикл, окружающий систему проводящих тканей, состоит из мелких живых тонкостенных клеток. Его толщина варьирует у разных растений. Многослойный перицикл встречается в корнях гинкго, саговника, принадлежащих к голосеменным растениям, а также в корнях грецкого ореха, хмеля, агавы, драцены из покрытосеменных. Некоторые хвойные имеют двуслойный перицикл, у большинства растений он однослойный (рис. 164 Б; 165). У злаков он не развивается против тяжей первичной ксилемы. У некоторых растений перицикл имеет неоднородное строение из-за развития в нем вместилищ выделений. Они характерны для некоторых хвойных, а из покрытосеменных — для представителей семейств зонтичных, аралиевых. У этих растений в перицикле, обычно против тяжей ксилемы, образуются схизогенные вместилища выделений.
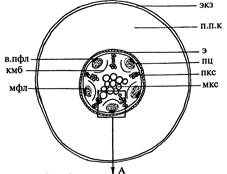
| Рис. 167. Заложение пучкового камбия в корне фасоли: А — схема первичного строения корня; Б — фрагмент клеточного строения. Обозначения: в.пфл — волокна протофлоэмы, кмб — камбий, мкс — метаксилема, п.К — перерезанный поясок Каспари, пкс — проток-силема, п.п.к — паренхима первичной коры, пц — перицикл, с.к — сопровождающая клетка, с.т — ситовидная трубка, э — эндодерма, экз — экзодерма | |||

| ||||
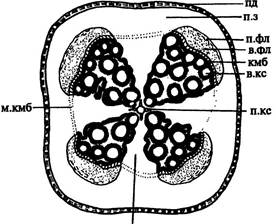
| В старых корнях однодольных все или отдельные группы клеток перицикла могут одревесневать, такой перицикл выполняет механическую роль. Анатомическое строение корня тесно связано с поглощением из почвы воды, которая должна кратчайшим путем достичь проводящей системы. и первичной корой, клетки которой постепенно отмирают; омертвевшая первичная кора, включая эндодерму, не выдерживающая давления разрастающихся проводящих тканей и перидермы, местами разрывается (рис. 168 Г) и впоследствии опадает. После удаления первичной коры покровной тканью корня становится пробка. Корни со слабым вторичным утолщением могут сохранять первичную кору продолжительное время, как у воронца, вьюнка, лютика. | |||
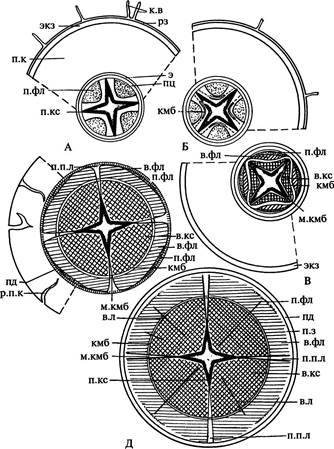
| Рис. 168. Стадии вторичного утолщения корня двудольного растения (А-Д). Обозначения: в.кс — вторичная ксилема, в.л — вторичный флоэмно-ксилемный луч, в.фл — вторичная флоэма, к.в — корневые волоски, кмб — пучковый камбий, м.кмб — межпучковый камбий, пд — перидерма, п.з — паренхимная зона, п.к — первичная кора, п.кс — первичная ксилема, п.п.л — первичный паренхимный луч, п.фл — первичная флоэма, пц — перицикл, рз — ризодерма, р.п.к — разрыв первичной коры, э — эндодерма, экз — экзодерма | ||||
 п.пл
п.пл
| Рис. 169. Вторичное строение корня тыквы (схема). Обозначения: в.кс — вторичная ксилема, в.фл — вторичная флоэма, кмб — пучковый камбий, м.кмб — межпучковый камбий, пд — перидерма, п.з — паренхимная зона, п.кс — первичная ксилема, п.пл — первичный паренхимный луч, п.фл — первичная флоэма | |||
Вторичные проводящие ткани в корнях состоят из тех же элементов, что и в стеблях.
Годовые приросты древесины в корнях деревьев тоньше, чем в стволах и ветвях, границы между ними выражены нечетко. В корнях хвойных трахеиды более крупные, чем в надземных органах, их стенки богаче порами, которые нередко расположены в несколько рядов. В корнях лиственных пород трахеи и трахеиды тонкостенные, широкопросветные и более многочисленные, распределены они равномернее, чем в стволах и многолетних ветвях. В корнях всегда больше тяжевой паренхимы и меньше либрифор-ма. Лучи не строго радиальные, тиллы образуются значительно реже, чем в стволовой древесине, типичная ядровая древесина бывает редко.
| 
| По степени развития механических элементов в лубе, толщине перидермы корни уступают надземным органам. Эти различия в строении осевых органов объясняются функциональными особенностями, так как стебель приспособлен к поддержанию обширной Таким образом, в процессе корнеобразования перицикл играет важнейшую роль. С возрастом он утрачивает эту способность и у некоторых однодольных, как уже было отмечено (гл. 10, п. 3.6), может склерифициро-ваться, превращаясь в механическую ткань. Типичные боковые корни закладываются в перицикле при первичном строении главного корня. У голосеменных и двудольных покрытосеменных растений новые корни развиваются и на поверхности старых, вторично утолщенных корней. Эти корни могут закладываться в камбиальной зоне, в месте ее пересечения лубо-древесинными лучами, как у яблони, тополя, в феллогене, как у щавелька, Иван-чая. Часто они развиваются около придаточных почек или в других местах, где имеются живые ткани. |
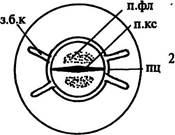
| 
| |

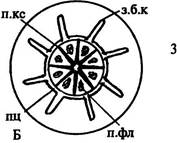
| 
| |
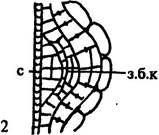
| Рис. 170. Начальные стадии развития (А) и места заложения боковых корней (Б). Обозначения: з.б.к — зачатки боковых корней, к.км — корневой кармашек, п.к — первичная кора, п.кс — первичная ксилема, п.п.к — паренхимные клетки первичной коры, п.пц — производные перицикла, п.фл — первичная флоэма, пц — перицикл, с — сосуд первичной ксилемы, э — эндодерма |
Так как эти корни нельзя называть боковыми, которые закладываются только в перицикле, Н. С. Воронин относит их к категории придаточных корней на корнях, противопоставляя их придаточным корням, образующимся на других органах растений.
Стеблевые придаточные корни могут закладываться и эндогенно, и экзо-генно. Начало этим корням дают клетки сердцевинных лучей, камбиальной зоны, флоэмы, феллогена, феллодермы и других живых тканей. Места их заложения нередко варьируют даже у одного и того же растения. Првдаточный корень, растущий в глубине материнского органа, также окружен корневым кармашком.
Развитие постоянных тканей и соединение проводящих систем придаточного корня и несущего его органа имеет много общего с развитием боковых корней.
Корни способны образовывать не только корни, но и придаточные почки, что определяет их участие в вегетативном размножении. Придаточные почки могут быть экзогенными и эндогенными. В первом случае они обычно закладываются в феллогене, феллодерме или наружных слоях первичной коры, во втором случае — в перицикле, камбиальной зоне, в разрастающихся лубо-древесинных лучах и других тканях стебля. Соединение проводящих систем почки и корня происходит аналогично соединению проводящих систем главного и боковых корней.
|
|
|
|
|
Дата добавления: 2015-05-08; Просмотров: 1423; Нарушение авторских прав?; Мы поможем в написании вашей работы!