КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Влияние запаздывания
|
|
|
|
Уравнения, которые представлены выше, предполагают, что процессы размножения и гибели происходят одновременно и популяция мгновенно реагирует на любое изменение внешних условий. Однако в реальности это не так. Всегда имеется некоторое запаздывание, которое вызвано несколькими причинами.
Развитие любой взрослой особи из оплодотворенного яйца требует определенного времени T. Поэтому если какое-нибудь изменение в окружающей среде, например, увеличение ресурса, вызовет внезапное повышение продуктивности взрослых особей, то соответствующе изменение численности произойдет лишь по прошествии времени T. Это означает, что уравнение:
dx/dt=f(x), (4)
где x - численность взрослых особей, следует заменить уравнением:
dx/dt=f(xt-T) (4.1)
где xt--T - численность половозрелых особей в момент t-T.
В реальных популяциях интенсивность размножения и гибели различны в разных возрастных группах. Например, у насекомых откладывают яйца взрослые особи, а конкуренция наиболее выражена на личиночной стадии. Такие процессы, как отравление среды продуктами метаболизма, каннибализм и т.п. в наиболее сильной степени воздействуют на ранние возрастные стадии, а их интенсивность зависит от численности взрослых особей, т.е. отрицательное влияние на коэффициент естественного прироста оказывают особи предыдущего поколения. С учетом этих обстоятельств, логистическое уравнение (3.1) перепишется в виде:
dx/dt=x(r-xt-T) (4.2)
Наиболее распространенное и изученное в динамике популяций уравнение Хатчинсона учитывает тот факт, что особи размножаются лишь с определенного возраста, и имеет вид:
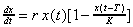 (4.3)
(4.3)
Смысл модели (4.3) заключается в том, что уровень лимитирования системы зависит не только от общей численности популяции в данный момент времени t, определяемой емкостью среды, но и от количества половозрелых особей в момент времени t-T. Еще более точное уравнение, учитывающее распределение времени запаздывания:
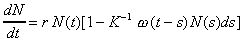
Вид функции распределения времен запаздывания w(t-s) представлен на рис. 4.
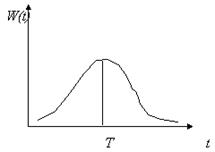
Рис.5. Модель динамики популяции с учетом распределения времен запаздывания. Типичный вид весовой функции w(t)
Такого типа уравнения могут иметь колебательные решения. Это легко проверить для простого линейного уравнения:
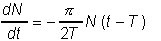 ,
,
которое имеет периодическое решение  в широком диапазоне значений скоростей роста r и времени запаздывания T.
в широком диапазоне значений скоростей роста r и времени запаздывания T.
Заключение
Популяционная динамика представляет собой область математической биологии, описывающая с помощью моделей типы динамическогоповедения развивающихся систем, представляющих собой одну или несколько взаимодействующих популяций или внутрипопуляционных групп. Отличительной чертой биологических популяций, как и всех живых систем, является их удаленность от равновесия, использование для своего роста и развития энергии внешних источников. Это обуславливает необходимость использования для описания таких систем нелинейных моделей, позволяющих отразить основные характерные черты популяционной динамики лабораторных и природных популяций. Это - ограниченность роста, вызванная совокупностью факторов. Возможность нескольких стационарных исходов в зависимости от начальных условий роста популяции. "Зависание" системы вблизи критической границы и ее чувствительность в этой области к малым флуктуациям и индивидуальным усилиям. Запаздывание реакции системы на изменение внешних факторов. Возможность колебательных и квазистохастических режимов.. Математические результаты, полученные при изучении моделей популяционной динамики служат для практических целей управления биотехнологическими и природными системами и, дают пищу для развития собственно математических теорий.
Список литературы
1. А.Д.Базыкин. Математичесакая биофизика взаимодействующих популяций. М., Наука, 1985, 165 с.
2. М.Бигон, Дж.Харпер., К. Таунсенд. Экология. Особи, популяции и сообщества. М., Мир. 1989, Том 1, 657 с.
3. Пайтген Х.-О., Рихтер П.Х. Красота фракталов. Образы комплексных динамических систем. М., Мир, 1993, 176 с.
4. Ризниченко Г.Ю., Рубин А.Б. Математические модели биологических продукционных процессов. М., Изд. МГУ, 1993, 301 с..
5. Ю.М.Свирежев, О.Д.Логофет. Устойчивость биологических сообществ. М., Наука, 1978, 352 с.
6. С.П.Капица, С.П.Курдюмов, Г.Г.Малинецкий. Синергетика и прогнозы будущего. М., Наука, 1997.
|
|
|
|
|
Дата добавления: 2015-05-10; Просмотров: 561; Нарушение авторских прав?; Мы поможем в написании вашей работы!