КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Влияние света на рост растений – фотоморфогенез. Влияние красного и дальнего
|
|
|
|
2.
3.Адаптация растений к отрицательныим температурам. Закаливание растений. Устойчивость растений к низким температурам подразделяют на холодостойкость и морозоустойчивость. Под холодостойкостью понимают способность растений переносить положительные температуры несколько выше 0 оС. Для характеристики холодостойкости растений используют понятие «температурный минимум», при котором рост растений прекращается. О холодостойкости растений косвенно можно судить по показателю суммы биологических температур. Чем меньше эта величина, тем быстрее растения созревают и тем выше их устойчивость к холоду. Показатели суммы биологических температур соответствуют скороспелости растений: очень раннеспелые имеют сумму биологических температур 1200 оС, раннеспелые - 1200-1600, среднераннеспелые – 1600 - 2200, среднеспелые – 2200 - 2800, среднепозднеспелые – 2800 - 3400, позднеспелые – 3400 - 4000 оС.
Холодостойкость теплолюбивых растений можно повысить двумя основными способами:
1) прививкой теплолюбивых растений на более холодоустойчивые подвои,
2) закаливанием.
Морозоустойчивость - способность растений переносить температуру ниже 0 °С.
Морозоустойчивость - не постоянное свойство растений. Она зависит от генетической природы организма, а также от физиологического состояния растения и условий внешней среды. Так, растения, выращенные при относительно низких положительных температурах, более устойчивы, чем выращенные при относительно высоких, в то же время один и тот же растительный организм в разных условиях способен погибнуть при температуре -5°С или перенести температуру до -40°С.
Другими словами, морозоустойчивость формируется в процессе всего развития (онтогенеза) растительного организма под влиянием определенных условий среды в соответствии с генотипом растения.
Морозоустойчивость некоторых растений (в частности, многих сортов роз) можно повысить комплексом мероприятий, и важное место среди них занимает закаливание.
Закаливание — это обратимое физиологическое приспособление растений к неблагоприятным воздействиям, происходящее под влиянием некоторых внешних условий.
Разные органы растений имеют неодинаковую способность к закаливанию, например, листья листопадных растений не обладают способностью к закаливанию; цветочные почки закаливаются хуже, чем листовые.
Процесс закаливания возможен лишь на определенных этапах развития растений. Для приобретения способности к закаливанию растения должны, прежде всего, закончить процессы роста. Сигналом к прекращению роста и стимулом для изменений в гормональной системе для растений является сокращение фотопериода и снижение температуры.
Под воздействием этих условий ослабляется синтез ростовых веществ – индолилуксусной кислоты и гиббереллинов – и усиливается образование абсцизовой кислоты и этилена. Это и приводит к торможению ростовых процессов.
Если в течение лета у многолетников процессы роста не успели закончиться, то это может вызвать массовую гибель растений зимой. Так, зимняя гибель часто вызывается летней засухой. Засуха приостанавливает рост летом, не позволяет многолетним культурам завершить ростовые процессы к осени. В результате растения оказываются неспособными пройти процессы закаливания и гибнут даже при небольших морозах. Аналогичная картина характерна для растений, выращенных при несоответствующем фотопериоде, не успевших завершить летний рост и поэтому неспособных к закаливанию. У вегетирующих растений легко вымерзают растущие и не закончившие рост органы.
Другим условием для приобретения способности к закаливанию является завершение оттока веществ. Всякое нарушение процессов оттока (кольцевание, например) препятствует закаливанию. Роль корней не сводится только к тому, что туда оттекают продукты обмена, гормоны, способствующие ростовым процессам. Важное значение имеет то, что клетки корня вырабатывают и накапливают вещества, повышающие устойчивость организма против мороза (криопротекторы).
Таким образом, процесс закаливания требует комплекса внешних условий и проходит в две фазы.
Первая фаза закаливания. Закаливание проходит на свету при пониженных плюсовых температурах (днем около +10, ночью около +2°С) и умеренной влажности. В этот период продолжается дальнейшее замедление и даже полная остановка ростовых процессов, а также с достаточной интенсивностью идет накопление криопротекторов, в частности образование сахаров.
Чем больше морозостойкость растения, тем большей способностью к накоплению сахаров в процессе фотосинтеза и к накоплению их именно при пониженной температуре оно обладает. Пониженная температура сокращает трату сахаров как в процессе дыхания, так и в процессах роста.
Влияние сахаров на повышение морозоустойчивости растений многосторонне. Накапливаясь в клетках, сахара повышают концентрацию клеточного сока, снижают водный потенциал. Чем выше концентрация раствора, тем ниже его точка замерзания, поэтому накопление сахаров стабилизирует клеточные структуры, в частности хлоропласты, благодаря чему они продолжают функционировать. Процесс запасания солнечной энергии в клетках продолжается даже при отрицательных температурах.
Особенное значение имеет защитное влияние сахара на белки, сосредоточенные в поверхностных мембранах клетки. Защитное действие сахаров проявляется только в том случае, если происходит при понижении температуры. Имеются данные, что сахара повышают устойчивость именно специфических белков, образующихся при пониженной температуре.
В первую фазу закаливания происходит также уменьшение содержания свободной воды. Излишняя влажность почвы (дождливая осень) препятствует прохождению процесса закаливания. Чем меньше в клетках и тканях содержание воды, тем меньше образуется льда и тем меньше опасность повреждения.
Влияние света в первую фазу закаливания не ограничивается увеличением накопления сахаров, помимо этого свет оказывает регуляторное воздействие. Это подтверждается тем, что этиолированные (обесцвеченные) растения не способны к закаливанию даже при обогащении их сахарами.
Среди механизмов адаптации к действию пониженных температур — синтез ряда стрессовых белков. Эти гидрофильные белки синтезируются в цитоплазме под действием низких температур и выделяются в клеточную стенку. Они располагаются на поверхности кристаллов льда, препятствуют их росту, тормозят образование межклеточного льда. Кроме того, образование таких белков позволяет использовать энергию окисления на поддержание температуры органов растений на 4—7 °С выше температуры окружающего воздуха.
К концу первой фазы закаливания клетки растений переходят в покоящееся состояние. Происходит процесс обособления цитоплазмы, что, в свою очередь, снижает возможность ее повреждения образующимися в межклетниках кристаллами льда. Особенно интенсивно перестройка обмена веществ протекает в период второй фазы закаливания.
Вторая фаза закаливания протекает при дальнейшем понижении температуры и не требует света. В связи с этим она может протекать и под снегом.
В эту фазу происходит отток воды из клеток, а также перестройка структуры протопласта. Продолжается новообразование специфических, устойчивых к обезвоживанию белков. Перестройка цитоплазмы способствует увеличению ее проницаемости для воды. Благодаря более быстрому оттоку воды уменьшается опасность внутриклеточного льдообразования.
Не для всех растений необходимо протекание процессов закаливания в две фазы. Например, у древесных растений, обладающих достаточным количеством сахаров, сразу протекают изменения, соответствующие второй.
Таким образом, в процессе закаливания возникает морозоустойчивость, которая определяется рядом изменений. У закаленных растений благодаря высокой концентрации клеточного сока, уменьшению содержания воды кристаллы льда образуются не в клетке, а в межклетниках. Количество образовавшегося льда в межклетниках у закаленных растении также значительно меньше. Изменение свойств белков цитоплазмы приводит к тому, что они становятся более устойчивыми к обезвоживанию. Накопление сахаров оказывает дополнительное защитное влияние. Цитоплазма закаленных растений более устойчива и к механическому давлению.
Билет 16. Физические свойства хлорофилла
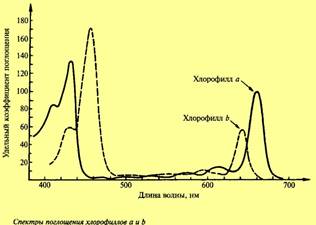
Как уже отмечалось, хлорофилл способен к избирательному поглощению света Спектр поглощения данного соединения определяется его способностью поглощать свет определенной длины волны (определенного цвета). Для того чтобы получить спектр поглощения, К.А. Тимирязев пропускал луч света через раствор хлорофилла. Часть лучей поглощалась хлорофиллом, и при последующем пропускании через призму в спектре обнаруживались черные полосы. Было показано, что хлорофилл в той же концентрации, как в листе, имеет две основные линии поглощения в красных и сине-фиолетовых лучах. При этом хлорофилл а в растворе имеет максимум поглощения 429 и 660 нм, тогда как хлорофилл b — 453 и 642 нм. Однако необходимо учитывать, что в листе спектры поглощения хлорофилла меняются в зависимости от его состояния, степени агрегации, адсорбции на определенных белках. В настоящее время показано, что есть формы хлорофилла, поглощающие свет с длиной волны 700, 710 и даже 720 нм. Эти формы хлорофилла, поглощающие свет с большой длиной волны, имеют особенно важное значение в процессе фотосинтеза.
Хлорофилл обладает способностью к флуоресценции. Флуоресценция представляет собой свечение тел, возбуждаемое освещением и продолжающееся очень короткий промежуток времени (10-8—10-9 с). Свет, испускаемый при флюоресценции, имеет всегда большую длину волны по сравнению с поглощенным. Это связано с тем, что часть поглощенной энергии выделяется в виде тепла. Хлорофилл обладает красной флуоресценцией.
Помимо фотосинтеза для растений, выращиваемых методом светокультуры, важное значение имеют процессы фотоморфогенеза, т. е. изменения размеров и формы под влиянием излучения разного качества и разной интенсивности.
Материальной основой, с помощью которой осуществляется формативное действие, служат, видимо, пигменты. Один из них – хромопротеид фитохром (фх). Установлено, что фитохром существует в двух формах: одна поглощает красное (red) излучение с длиной волны 660 нм (Фк), а другая — дальнее красное (far-red) с длиной волны 730 нм (Флк). Под действием красного излучения Фк активизируется и переходит в Фдк, а под действием дальнего красного – наоборот. Установлено, что фитохром регулирует прорастание семян, растяжение стеблей, образование листьев и пигментов, формирование корневой системы и некоторые другие физиологические процессы, ход которых определяется наличием взаимоисключающих влияний красного или дальнего красного излучения (red-far-red effect).
Установлено также сильное формативное действие синего излучения с длиной волны 450 нм. Это излучение избирательно поглощается каротиноидами, флавинами и пластохинонами.
Формативное действие оптического излучения проявляется также в виде:
фототропизма, т. е. неравномерного роста вследствие одностороннего облучения;
фототаксиса – движения как в сторону источника, так и от него;
фотонастий — ненаправленных кратковременных движений.
Эти процессы редко оказывают значительное влияние на рост и урожай сельскохозяйственных культур. Более подробно они изложены в курсе физиологии растений.
Фотопериодизм сельскохозяйственных культур изучен еще недостаточно. Известно, что отдельные сорта томатов или огурцов неодинаково реагируют на разное соотношение светлого и темного периода суток. В летнее время эти культуры успешно растут и плодоносят и на юге (Крым), и на Крайнем Севере (Кольский п-ов).
Из курса физиологии растений известно, что фотопериодическая реакция зависит от качества излучения, температуры воздуха, этапа развития растений и многих других факторов.
|
|
|
|
|
Дата добавления: 2015-06-25; Просмотров: 1853; Нарушение авторских прав?; Мы поможем в написании вашей работы!