КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Фикобилины и их роль в фотосинтезе
|
|
|
|
Фикобилины — красные и синие пигменты, содержащиеся у цианобактерий и некоторых водорослей. Исследования показали, что красные водоросли и цианобактерий наряду с хлорофиллом а содержат фикобилины. В основе химического строения фикобилинов лежат четыре пиррольные группировки. В отличие от хлорофилла у фикобилинов пиррольные группы расположены в виде открытой цепочки. Фикобилины представлены пигментами: фикоцианином, фикоэритрином и аллофикоцианином. Фикоэритрин — это окисленный фикоцианин. Красные водоросли в основном содержат фикоэритрин, а цианобактерий — фикоцианин. Фикобилины образуют прочные соединения с белками (фикобилинпротеиды). Связь между фикобилинами и белками разрушается только кислотой. Предполагается, что карбоксильные группы пигмента связываются с аминогруппами белка. Необходимо отметить, что в отличие от хлорофиллов и каротиноидов, расположенных в мембранах, фикобилины концентрируются в особых гранулах (фикобилисомах), тесно связанных с мембранами тилакоидов.
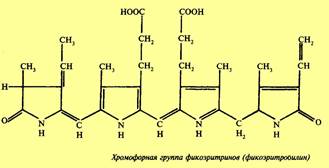
Хромофорная группа фикоэритринов (фикоэритробилин)
Фикобилины поглощают лучи в зеленой и желтой частях солнечного спектра. Это та часть спектра, которая находится между двумя основными линиями поглощения хлорофилла. Фикоэритрин поглощает лучи с длиной волны 495— 565 нм, а фикоцианин — 550— 615 нм. Сравнение спектров поглощения фикобилинов со спектральным составом света, в котором проходит фотосинтез у цианобактерий и красных водорослей, показывает, что они очень близки. Это позволяет считать, что фикобилины поглощают энергию света и, подобно каротиноидам, передают ее на молекулу хлорофилла, после чего она используется в процессе фотосинтеза. Наличие фикобилинов у водорослей является примером приспособления организмов в процессе эволюции к использованию участков солнечного спектра, которые проникают сквозь толщу морской воды (хроматическая адаптация). Как известно, красные лучи, соответствующие основной линии поглощения хлорофилла, поглощаются, проходя через толщу воды. Наиболее глубоко проникают зеленые лучи, которые поглощаются не хлорофиллом, а фикобилинами.
2 Движение растений.
Тропизмы, настии. Растительный организм обладает способностью к определенной ориентировке своих органов в пространстве. Реагируя на внешние воздействия, растения меняют ориентировку органов. Различают движения отдельных органов растения, связанные с ростом — ростовые и с изменениями в тургорном напряжении отдельных клеток и тканей — тургорные. Ростовые движения, в свою очередь, бывают двух типов: тропические движения, или тропизмы,— движения, вызванные односторонним воздействием какого-либо фактора внешней среды (света, силы земного притяжения и др.); настические движения, или настии,— движения, вызванные общим диффузным изменением какого-либо фактора (света, температуры и др.). В зависимости от фактора, вызывающего тропические движения, различают геотропизм, фототропизм, хемотропизм, тигмотропизм, гидротропизм. Геотропизм — движения, вызванные односторонним влиянием силы тяжести. Если положить проросток горизонтально, то через определенный промежуток времени корень изгибается вниз, а стебель — вверх. Еще в начале ХIХ в. был изобретен прибор клиностат. В этом приборе проросток в горизонтальном положении привязывается к вращающейся оси. Благодаря этому сила притяжения действует попеременно на нижнюю и верхнюю стороны проростка. В этом случае рост проростка идет строго горизонтально и никаких изгибов не наблюдается. Эти опыты доказали, что изгибы стебля и корня связаны с односторонним действием силы земного притяжения. Изгиб корня вниз (по направлению действия силы притяжения) называют положительным геотропизмом.
Тропизмы и настии: 1 — геотропизм; 2 — фототропизм; 3 — термонастии; 4 — фотонастии.
Геотропическая реакция — «пороговое» явление, т. е. геотропический изгиб происходит лишь при достижении раздражителем какого-то определенного уровня. Количество раздражителя равно силе гравитации, умноженной на время. Для того чтобы произошел изгиб, проросток должен быть выдержан в горизонтальном положении определенное время (время презентации). Если проростки выдержать это время в горизонтальном положении, а затем поместить вертикально, то все равно изгиб произойдет. При строго горизонтальном положении проростка время презентации наименьшее. Чем ближе положение проростка к вертикальному, тем больше время презентации. Это понятно, так как сила земного притяжения наибольшая при горизонтальном положении. Время презентации составляет примерно 3—5 мин. Время, необходимое для проявления изгиба, — 45—60 мин. Направление геотропической реакции может изменяться в процессе роста организма, а также в зависимости от условий среды. Так, для цветоножки мака до распускания бутона характерен положительный геотропизм, а после распускания цветков — отрицательный. При пониженной температуре отрицательный геотропизм стебля может переходить в диагеотропизм (стелющиеся формы).
Фототропизм — движения, вызванные неравномерным освещением разных сторон органа. Если свет падает с одной стороны, стебель изгибается по направлению к свету — положительный фототропизм. Корни обычно изгибаются в направлении от света — отрицательный фототропизм. Ориентировку пластинок листьев перпендикулярно к падающему свету (при большой интенсивности света) называют диафототропизмом. Для восприятия одностороннего освещения также необходимо определенное время презентации, которое зависит от силы одностороннего освещения. В зависимости от возраста растения и от условий среды направление фототропических изгибов может меняться. Так, у настурции до цветения для стебля характерен положительный фототропизм, а после созревания семян — отрицательный.
Хемотропизм — это изгибы, связанные с односторонним воздействием химических веществ. Хемотропические изгибы характерны для пыльцевых трубок и для корней растений. Если пыльцу положить на предметное стекло в среду, содержащую сахарозу, и одновременно поместить туда кусочек завязи, все пыльцевые трубки в процессе роста изогнутся по направлению к завязи. Корни растений изгибаются по направлению к питательным веществам. Если питательные вещества не перемешаны со всей почвой, а распределяются отдельными очагами, корни растут по направлению к этим очагам. Такая способность корней определяет большую эффективность гранулированных удобрений. Корни растут по направлению к отдельным гранулам, содержащим питательные вещества. При таком способе внесения питательных веществ создается также повышенная концентрация их около корня, что обусловливает их лучшую усвояемость.
Гидротропизм — это изгибы, происходящие при неравномерном распределении воды. Для корневых систем характерен положительный гидротропизм.
Аэротропизм — ориентировка в пространстве, связанная с неравномерным распределением кислорода. Аэротропизм свойствен в основном корневым системам.
Тигмотропизм — реакция растений на одностороннее механическое воздействие. Тигмотропизм свойствен лазающим и вьющимся растениям.
Настические движения бывают двух типов: эпинастии — изгиб вниз и гипонастии — изгиб вверх. В зависимости от фактора, вызывающего те или иные настические движения, различают термонастии, фотонастии, никтинастии и др.
Термонастии — движения, вызванные сменой температуры. Ряд растений (тюльпаны, крокусы) открывают и закрывают цветки в зависимости от температуры. При повышении температуры цветки раскрываются (эпинастические движения), при снижении температуры закрываются (гипонастические движения).
Фотонастии — движения, вызванные сменой света и темноты. Цветки одних растений (соцветия одуванчика) закрываются при наступлении темноты и открываются на свету. Цветки других растений (табака) открываются с наступлением темноты.
Никтинастии («никти» — ночь) — движения цветков и листьев растений, связанные с комбинированным изменением, как света, так и температуры. Такое комбинированное воздействие наступает при смене дня ночью. Примером являются движения листьев у некоторых бобовых, а также у кислицы. К ростовым движениям относятся и круговые движения концов молодых побегов и кончиков корней относительно оси. Такие движения называют круговые нутации. Примерами являются движения стеблей вьющихся растений (хмель), усиков лазящих растений. Это необходимо для поиска опоры при движении к свету стеблей.
Тургорные движения. Не все настические движения относятся к ростовым. Некоторые связаны с изменением тургора. К ним относятся никтинастические движения листьев. Так, для листьев многих растений характерны ритмические движения — у клевера наблюдается поднятие и складывание листочков сложного листа ночью. Этот тип движений связан с изменением тургора в специализированных клетках листовых подушечек.
Сейсмонастии — движения, вызванные толчком или прикосновением, например движение листьев у венериной мухоловки или у стыдливой мимозы. В результате прикосновения листья мимозы опускаются, а листочки складываются. Реакция происходит чрезвычайно быстро, спустя всего 0,1 с. При этом раздражение распространяется со скоростью 40—50 см/с. Сейсмонастические движения листьев мимозы могут происходить и под влиянием местных тепловых, электрических или химических воздействий. Эти движения связаны с потерей тургора нижней стороны листа. Значение подобной двигательной реакции заключается в предотвращении повреждений от ливневых дождей и сильных ветров.
Автонастии — самопроизвольные ритмические движения листьев, не связанные с какими-либо изменениями внешних условий. Так, листья тропического растения десмидиум претерпевают ритмические колебания.
3. Роль изменения свойств мембран при воздействии неблагоприятных факторов на
растение. Отличительные особенности строения мембран различных экотипов растений.
Важнейшей неспецифической реакцией на неблагоприятные воздействия является изменение свойств мембран, что связано с перестройками в их структуре. Это в значительной мере касается липидов. Наблюдаются сдвиги в соотношении различных групп жирных кислот, изменяется степень их ненасыщенности, возрастает уровень перекисного окисления липидов (ПОЛ), снижается их подвижность. Это влияет на функции мембранных белков. Структурные изменения в мембранах приводят к освобождению из связанного состояния ионов Са2+. Известна роль кальция в поддержании структуры хроматина, в регуляции активности ферментов в митохондриях и хлоропластах. В цитозоле концентрация кальция невысока (10-5—10-8 М), в то время как в апопласте и органеллах его в 103—104 раз выше. В результате стрессового воздействия поток кальция из апопласта в цитоплазму резко возрастает. Вслед за этим кальций выводится из цитоплазмы. Изменение концентрации кальция запускает специфические мембранные каналы и транспортные системы, а также вызывает структурные изменения в клетке. Нарушение структуры мембран приводит к многочисленным изменениям в метаболизме. Повышается проницаемость мембран, происходит деполяризация мембранного потенциала плазмалеммы, значение рН сдвигается в кислую сторону. Возрастает активность Н+ — помпы в плазмалемме и тонопласте. Увеличивается вязкость цитоплазмы, наблюдается торможение деления и роста клеток. Важной особенностью реакции растений на стресс-факторы является изменение напряженности энергетического обмена. Митохондрии являются основными органеллами, снабжающими клетки таким энергетическим эквивалентом как АТФ. В состоянии стресса цитохромный путь дыхания падает и возрастает альтернативный путь с его терминальной оксидазой АО, не сопровождаемой образованием АТФ. Возникает недостаток энергетических ресурсов. Между тем при стрессе необходимы дополнительные энергетические эквиваленты. Возрастают затраты АТФ на поддержание структуры и обменавеществ, что сопровождается временной активацией дыхания. В дальнейшем при усилении действия стрессора дыхание снижается, и соотношение синтеза и расхода АТФ еще больше нарушается. Возрастание активности гидролитических процессов ведет и к накоплению различных протекторных соединений, например такого низкомолекулярного осмотически активного вещества, как пролин. Пролин способен образовывать гидрофильные коллоиды, что удерживает воду и защищает белки от денатурации (при засухе, засолении, низкой или высокой температурах). Из других стрессовых метаболитов необходимо отметить образование редуцирующих Сахаров, полиаминов, бетаинов. Полиамины способны предотвращать повреждения, вызванные морозом, засухой, действием солей. Это вещества основной природы, легко связываются с отрицательно заряженными группами полимеров. Увеличивают стабильность РНК, ДНК, рибосом, стабилизируют мембраны, тормозят лизис клеточных стенок. Бетаины как метилированные производные аминокислот и аминов являются главными донорами метильных групп. Метилирование изменяет функциональную активность ДНК и других внутриклеточных полимеров и повышает их устойчивость к различным стрессовым воздействиям. Следует отметить, что перенос метильных групп на ДНК является самой распространенной модификацией ДНК. Существенную роль в ответе растений на стрессоры играет гормональная система. Показано, что при неблагоприятных условиях возрастает количество абсцизовой кислоты, этилена, жасмоновой кислоты, изменяется соотношение фитогормонов. Высказывается мнение, что у растений в отличие от животных при неблагоприятных условиях ведущую роль играют гормоны, тормозящие их функциональную активность. Это обеспечивает организму торможение роста и вхождение в покоящееся состояние. При действии неблагоприятных факторов важным для растения, является соранение нормальных донорно-акцепторных отношений, поскольку при снижении запроса на ассимиляты, уменьшается фотосинтез. Однако на уровне организма имеется возможность смягчать это явление. В ряде случаев это происходит в результате увеличения объемов запасающих тканей.
Устойчивость растений против неблагоприятных условий имеет разный характер. Она может быть основана на том, что организм тем или иным путем избегает их воздействия. Например, одни растения запасают воду (суккуленты) и тем самым избегают обезвоживания при засухе, другие растения, с очень коротким вегетационным периодом (эфемеры), приурочивают жизнедеятельность ко времени выпадения осадков. Значительно большее значение имеет устойчивость, основанная на выносливости клеток растений, т. е. способности в процессе адаптации перестраивать как скорость, так и направление метаболических реакций таким образом, чтобы и в изменившихся условиях среды вырабатывать все необходимые соединения. Разный уровень устойчивости обусловлен биологическими особенностями видов. Растения, различающиеся по устойчивости, на стрессовые воздействия реагируют однотипно, но отличаются по скорости физиологических и структурных перестроек. Для растений, устойчивых к действию стрессоров, показана большая стабильность клеточных мембран по сравнению с неустойчивыми. Сохранению целостности мембран способствует торможение распада липидов и белков, что может быть связано с эффективной работой механизмов антиоксидантной защиты. Важными является сохранение содержания ненасыщенных жирных кислот, регуляция кальциевого обмена в клетках. Это придает мембране большую пластичность и создает лучшие условия для функционирования мембран. Так, показано, что мембраны холодостойких растений отличаются большим содержанием ненасыщенных жирных кислот. Это позволяет сохранять жидкостное состояние при пониженных температурах, при которых они переходят в полужидкое состояние. Значение липидов мембран в устойчивости к низким температурам подтверждается опытами с трансгенными растениями. При изучении процессов устойчивости отмечаются случаи одновременного ее повышения к нескольким факторам после действия какого-либо одного стрессора (по П.А. Генкелю: «сопряженнаяустойчивость»). Так, установлено, что предварительный тепловой шок повышает устойчивость к водному дефициту, засолению, тяжелым металлам. С другой стороны, повышение теплоустойчивости растений отмечено при водном дефиците, действии засоления, холодовой закалке. Из неблагоприятных условий, которые вызывают стресс у растительных организмов, наиболее часто встречающимися стрессорами являются недостаток воды, высокая температура, низкая температура, высокая концентрация солей. В настоящей главе остановимся на рассмотрении физиологических основ засухоустойчивости, жаростойкости, холодоустойчивости, морозоустойчивости, зимостойкости, солеустойчивости, устойчивости к недостатку или отсутствию кислорода
|
|
|
|
|
Дата добавления: 2015-06-25; Просмотров: 5223; Нарушение авторских прав?; Мы поможем в написании вашей работы!