КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Биохимические особенности нервной ткани 6 страница
|
|
|
|
К таким явлениям относятся изменения параметров вызванных потенциалов коры головного мозга.
Вызванные потенциалы, впервые описанные Эдрианом (1936), а затем многими другими авторами — Жувэ (1957), Эрнандес-Пеоном (1961, 1966), Даусоном (1958, 1959), И.А.Пеймером (1957) и др., — как известно, представляют собой электрические ответы на различные (зрительные, слуховые и др.) раздражители в соответствующих (затылочных, височных и др.) областях мозговой коры и подкорковых структурах. Параметры вызванных потенциалов изменяются в зависимости от интенсивности раздражителя, с одной стороны, и активности субъекта — с другой, причем изменения параметров вызванных потенциалов могут распространяться на ранние фазы ответа (т.е. возникать в интервале 15 — 100 мс) или касаться более поздних фаз (от 100 мс до 250 — 300 мс и больше).
Параметры вызванных потенциалов могут быть использованы не только как индикатор относительно элементарной анализаторной деятельности мозга, они с успехом могут быть использованы и для объективной регистрации тех изменений в приеме и переработке информации, которые возникают при мобилизации активного внимания.
Изучение вызванных потенциалов как объективного индикатора внимания можно использовать двумя путями.
С одной стороны, можно изучать изменения вызванных потенциалов при отвлечении внимания побочным раздражителем, с другой стороны, — в условиях привлечения внимания к данному раздражителю. Первый вариант опыта был проведен в классических исследованиях Эрнандес-Пеона (1956, 1960, 1969), показавшего, что корковые звуковые потенциалы резко тормозятся при предъявлении кошке вида или запаха мыши. Второй вариант опыта был проведен преимущественно на людях и заключался в том, что вызванные потенциалы в ответ на сенсорные раздражители сравнивались с вызванными потенциалами, полученными в условиях активного ожидания этих сигналов (соответственно предупреждающей инструкции) или в условиях усложнения анализа этих раздражителей и т. п. Факты, полученные в этих условиях рядом исследователей (Линдсли, 1960, 1961; А.И.Пеймер, 1958, 1966; Э. Г. Симерницкая, 1970; Тесце, 1970; и др.), показали, что привлечение внимания активным ожиданием или усложнением задачи приводит к заметному повышению амплитуды поздних компонентов вызванных потенциалов по сравнению с «фоном». Таким образом, изменение параметров потенциалов (особенно поздних компонентов) является отчетливым объективным признаком селективного внимания.
Как показали многочисленные исследования, введение задания (например, переход к инструкции внимательно следить за изменениями сигнала) приводит не только к увеличению амплитуды вызванных потенциалов, но и к распространению на другие зоны коры головного мозга, лежащие вне пределов коркового «ядра» данного анализатора.
Пример такого вовлечения вторичных и третичных отделов коры по мере мобилизации внимания соответствующей инструкцией в вызванную активность можно видеть на рис. 101. Этот рисунок отчетливо показывает, что предварительная инструкция не только увеличивает амплитуду вызванного потенциала, но и ведет к его
1. Ожидание тока
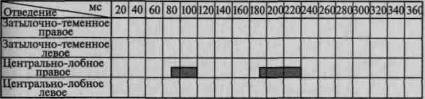
2. Дифференцирование зрительных стимулов
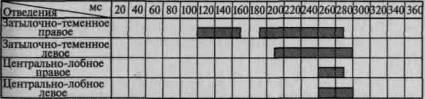
3.Выполнение произвольных двигательных реакций
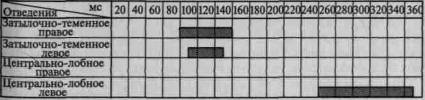
Рис. 101. Распространение достоверных изменений вызванных
потенциалов на отдельные зоны коры при активации избирательного
внимания (по Э. Г. Симерницкой)
распространению на другие более широкие области коры. Данные изменения параметров вызванных ответов при привлечении внимания взяты из исследования, проведенного в нашей лаборатории Э. Г. Симерницкой (1970).
Объективные электрофизиологические исследования позволяют не только изучать физиологические механизмы различных видов внимания; они дают также возможность проследить основные этапы их формирования. Эту возможность показали работы, проведенные Д. А. Фарбер и ее сотрудниками (Д.А. Фарбер, 1969; Е. М. Фрид, 1970; Д.А.Фарбер, Е.М.Фрид, 1971). Установлено, что если рефлекс вызывает заметное повышение амплитуды вызванных потенциалов даже у маленького ребенка, то стойкие изменения вызванных потенциалов в результате речевой инструкции (считать сигналы или наблюдать за их изменениями) носят у него еще недостаточно выраженный и устойчивый характер и начинают отчетливо и стойко выступать лишь на позднейших этапах развития ребенка.
Эти данные, приведенные на рисунках 102 и 103, показывают, что физиологические изменения, вызываемые речевой инструкцией и лежащие в основе произвольного внимания, формируются лишь к 12—15-летнему возрасту. Один из наиболее существенных для нас фактов заключается в том, что именно к этому возрасту выраженные и стойкие изменения в вызванных потенциалах начинают возникать не только в сенсорной области коры, но и в ее лобных отделах, и что лобные отделы коры, по всем данным, начинают к этому времени принимать ближайшее участие в обеспечении сложных и устойчивых форм высшего, произвольного внимания.
К специальному рассмотрению этого вопроса, представляющего большой нейропсихологический интерес, мы и обратимся.
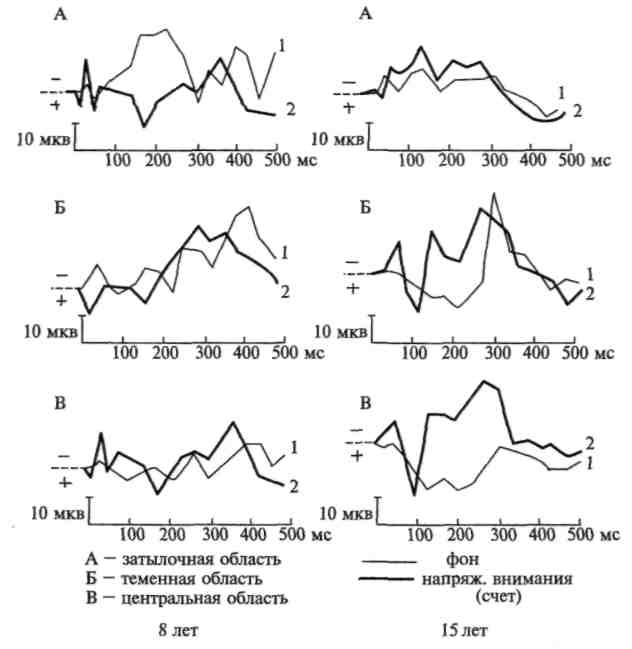
Рис. 102. Увеличение амплитуды вызванных потенциалов при напряжении активного внимания (по Д. А. Фарбер и Е. М. Фрид)
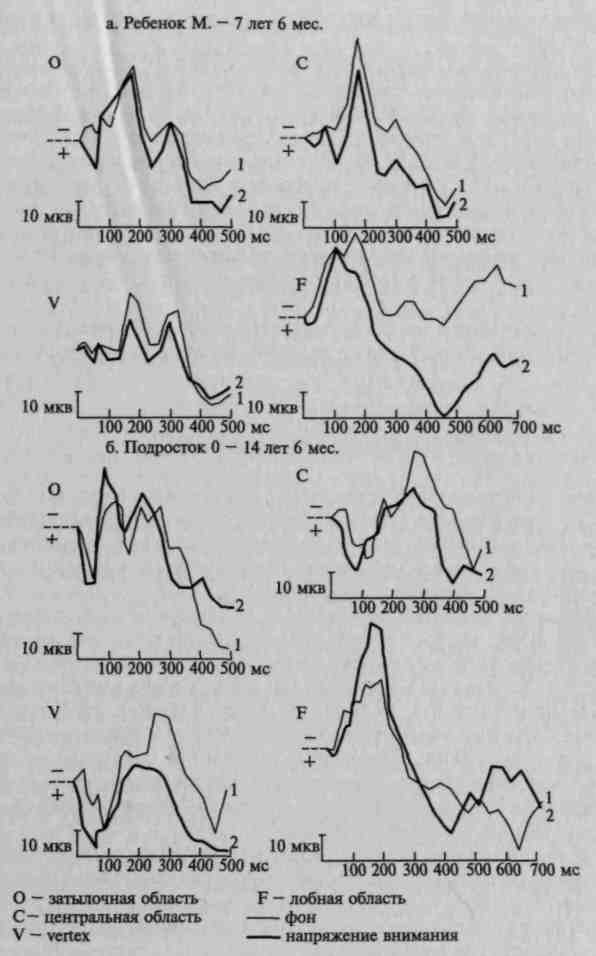
Рис. 103. Распространение изменений в вызванных потенциалах на
различные зоны коры при мобилизации активного внимания у детей
разного возраста. Изменение вызванных потенциалов под влиянием
речевой инструкции оценивать различные зрительные сигналы:
а — у ребенка 7 лет 6 мес; 6 — у подростка 14 лет 6 мес. (по Д. А. Фарбер и Е. М. Фрид)
3 МОЗГОВАЯ ОРГАНИЗАЦИЯ
Есть все основания считать, что не все системы переднего мозга играют одинаковую роль в процессах организации внимания и что вопрос о мозговой организации внимания следует дифференцировать как в отношении отдельных структур мозга, так и в отношении отдельных форм внимания.
Рассматривая вопрос об основных функциональных блоках головного мозга, мы уже указывали ранее на ту роль, которую играют образования верхних отделов ствола и ретикулярной формации среднего м о з г а в поддержании уровня бодрствования и в появлении общей реакции пробуждения (arousal). Именно поэтому после работ Мэгуна и Моруцци (1949), Линдсли и др. (1950), Джаспера (1956) данные образования считались основными механизмами, определяющими переход от сна к бодрствованию и обеспечивающими наиболее генерализованные и элементарные формы внимания.
Эти предположения подтвердились не только большим числом широко известных опытов с животными, при которых перерезка стволовой ретикулярной формации на определенном уровне приводила ко сну, а ее раздражение — к повышению бодрствования и обострению чувствительности (см. обзор этих фактов Линдсли, 1960), но и рядом клинических наблюдений, установивших, что поражения верхних отделов ствола и стенок третьего желудочка могут приводить к возникновению сна или к онейроидному, про-соночному состоянию и что тонус коры в этих случаях оказывается резко сниженным, а селективные формы сознания и внимания — глубоко нарушенными.
Однако стволовые и мезэнцефалические механизмы восходящей активирующей ретикулярной формации обеспечивают лишь наиболее генерализованные состояния бодрствования и внимания.
Для обеспечения избирательных (селективных) форм внимания, т. е. выделения того или другого сигнала и торможения реакций на побочные раздражители, необходимо участие более высоко расположенных образований головного мозга неспецифических ядер таламуса — лимбической коры и лобной области больших полушарий.
За последнее время накоплено много данных, освещающих ту роль, которую играют образования древней коры, или лимбической области (гиппокамп, миндалина), и связанные с ними аппараты хвостатого ядра в регуляции внимания и бодрствования. Исследования, проведенные на нейронном уровне, показали, что именно в этих образованиях (особенно в гиппокампе) значительное место занимают нейроны, отвечающие не на модально-специфические раздражения, а производящие как бы сличение (компарацию) старых и новых раздражителей и обеспечивающие реакцию на новые сигналы или их свойства с угашением реакции на старые, уже привычные раздражители (О.С.Виноградова, 1969, 1970). Поэтому образования гиппокампа, интимно участвующие в обеспечении механизмов торможения посторонних раздражителей и привыкании (habituatiori) к длительно повторяющимся раздражителям, стали считать одной из существенных составных частей того тормозящего или «фильтрующего» аппарата, который необходим для избирательных реакций на специфические раздражители, входящие в систему врожденных ориентировочных рефлексов и инстинктивного поведения. Именно в силу этого гиппокамп, а затем и хвостатое ядро стали рассматривать как существенные аппараты, обеспечивающие организму возможность осуществлять строго избирательные формы поведения (Граштиан, 1961; Дуглас, Прибрам, 1966; О.С.Виноградова, 1969, 1970), а нарушение этих образований — как источник распада селективности поведения в целом, которое в большей степени можно отнести к нарушениям избирательного внимания, чем к дефектам памяти. Этим представлениям о роли лимбических структур, и в частности образований гиппокампа, в организации поведения отвечают и те изменения в поведении и в состоянии сознания, которые наблюдаются у человека при поражениях этой области, а также при изучении в специальных электрофизиологических исследованиях.
Клинически больные этой группы — с расположенными по средней линии глубокими опухолями — не дают отчетливых нарушений гнозиса и праксиса, речи или формально-логических процессов. Патология психических процессов у данных больных заключается в выраженном нарушении избирательности, селективности психических процессов, в резкой их истощаемости.
В относительно более стертых случаях это проявляется в повышенной отвлекаемости больных, быстром прекращении активной, направленной деятельности, в легком всплывании побочных ассоциаций и в отчетливо выступающих нарушениях памяти, к которым мы еще обратимся далее. В более тяжелых случаях (массивные опухоли, влияющие на стенки третьего желудочка и лимбические структуры) этот синдром может принимать грубо выраженные формы и переходить в просоночные онейроидные состояния, при которых больные теряют грань между настоящим и прошлым, обнаруживают явления спутанности, конфабуляции. В обоих случаях любые посторонние раздражители сразу же вплетаются в течение мыслей, и организованное, избирательное строение сознания уступает место спутанности. Такие больные могут заявить, что они находятся не в больнице, а дома или на работе (а иногда и там, и здесь), что тут же рядом находятся их родные, они принимают врача за знакомого по месту работы и дают признаки грубых расстройств сознания, которые оказываются особенно отчетливо выраженными, если в патологический процесс вовлекаются передние отделы лимбической области и если критика к собственному состоянию нарушается (А. Р. Лурия, Е. Д.Хомская, С.М.Блинков, М.Кричли, 1967).
Описанные клинические факты нашли свое частичное разъяснение в тех электрофизиологических данных, которые были получены в последнее время при исследовании больных с массивными поражениями диэнцефальной области, опухолями стенок третьего желудочка и примыкающих к ним отделов лимбической системы.
В этих исследованиях (Л.П.Латаш, 1968; Е.Д.Хомская, 1969, 1972; Н.А.Филиппычева, Т.О.Фаллер, 1970; и др.) было отчетливо показано, что вегетативные и электрофизиологические компоненты ориентировочного рефлекса в этих случаях оказываются резко угнетенными, и даже повторные раздражители начинают вызывать лишь очень слабые и быстро угасающие реакции, что является отчетливым результатом дефицита тех неспецифических активирующих влияний, которые в норме обеспечивают соответствующий тонус коры мозга.
Закон силы, по которому сильные раздражители вызывают сильные, а слабые раздражители — слабые реакции, нарушается, разные по интенсивности раздражения начинают вызывать одинаковые слабые реакции, часто предъявленное раздражение вызывает не депрессию, а парадоксальную экзальтацию альфа-ритма. Процесс привыкания к новому раздражителю (habituation), отчетливо наблюдаемый при нормальном состоянии коры, также претерпевает грубые изменения, и электрофизиологические показатели ориентировочного рефлекса либо вообще не вызываются, либо становятся неугасимыми.
Одновременно эти исследования показали, что у данной группы больных можно наблюдать и известные извращения нормального протекания нервных процессов, что вплотную подводит нас к объяснению механизмов только что описанных психических явлений.
Существенным для этих случаев оказывается тот факт, что все эти признаки первичного нарушения селективного внимания, по данным ЭЭГ, могут в той или иной степени компенсироваться введением речевой инструкции, иначе говоря, включением в систему более сохранных высших уровней построения процесса. Чаще всего эта компенсация имеет лишь временный характер, и избирательность процессов активации быстро теряется; однако эта потенциальная сохранность высших, произвольных форм внимания при первичном нарушении его элементарных форм является одним из важных признаков, отличающих этих больных от больных с иной локализацией поражения (Е.Д.Хомская, 1972).
Совершенно иную роль в организации внимания играют лобные доли головного мозга.
В классических исследованиях над животными, как уже упоминалось ранее, считалось, что лобные доли мозга имеют непосредственное отношение к сохранению следов памяти, что и проявляется в факте нарушения отсроченных реакций после экстирпации лобной коры. Однако, как известно, этот факт получил в дальнейших работах совершенно иное истолкование (Малмо, 1942; Прибрам, 1959, 1963; Вейзкранц, 1968; и др.). Показано, что животное с удаленными лобными долями не могло сохранить отсроченных реакций не столько потому, что оно не удерживало прежних следов, а вследствие постоянного отвлечения побочными раздражителями. Устранение этих побочных раздражителей (например, путем помещения животного в полную темноту или проведение опытов на фоне транквилизаторов, снижающих общий тонус коры) приводило к восстановлению отсроченных реакций. Существенная роль лобных долей в торможении реакции на любые побочные раздражители и в сохранении направленного, программированного поведения была показана и опытами Конорского (1964; и др.), Брутковского (1964, 1966) и др. Именно в силу нарушения тормозящих механизмов экстирпация лобных долей неизменно приводила у животных к грубым нарушениям целенаправленного, избирательного поведения и к расторможению реакций на побочные раздражения. Соответствующие факты изменения поведения животных описаны в классических работах Бианки (1895, 1921), Джекобсена (1935), П. К.Анохина (1949), Прибрама (1954, 1958; и др.) и др.
Богатейшие данные о нарушениях избирательного поведения, и прежде всего высших произвольных форм внимания, дает клиника поражений лобных долей мозга.
В отличие от описанных случаев поражений верхних отделов ствола и лимбической области элементарные формы непроизвольного внимания или импульсивно возникающие ориентировочные реакции на любые раздражители могут быть у таких больных не только сохранены, но даже и патологически усилены; наоборот, всякие попытки вызвать у этих больных устойчивое произвольное внимание по речевой инструкции не приводят ни к каким результатам.
Невозможность сосредоточиться на данной инструкции и затормозить реакции на любые побочные раздражители становятся видны уже при обычных клинических наблюдениях над больными с массивными поражениями лобных долей мозга. Обычно такие больные начинают выполнять предложенное им задание, но лишь только в палату входит посторонний человек или сосед по палате обращается к кому-либо шепотом, больной тотчас же перестает выполнять начатое задание, переводит взор на вошедшего в палату или вмешивается в разговор соседа. Именно поэтому, чтобы исследовать такого больного, надо проводить опыт с его соседом: в этих случаях он непроизвольно вплетается в начатый разговор, обнаруживая при этом ту непроизвольную активность, которую трудно вызвать, обращаясь к нему с непосредственно адресованными ему заданиями.
Эта повышенная отвлекаемость больного с массивным поражением лобных долей становится источником глубоких нарушений его целенаправленного поведения.
Глубокое нарушение произвольного внимания или высших форм контроля над деятельностью при оживлении элементарных форм внимания и создает ту картину нарушений поведения, которую мы так часто встречаем в клинике поражений лобных долей мозга.
Ряд психофизиологических исследований, проведенных за последние годы, позволяет подойти к выделению механизмов, лежащих в основе нарушения высших произвольных форм внимания.
Как уже указывалось ранее (ч. вторая, гл. V), Грей Уолтер установил факт, что именно в лобных долях человека ожидание какого-либо сигнала вызывает специфические медленные потенциалы, названные им «волнами ожидания», и что лишь впоследствии эти волны распространяются на другие участки мозга (рис. 48). По данным М.Н.Ливанова и его сотрудников, каждое интеллектуальное напряжение вызывает в лобных отделах коры закономерное повышение числа синхронно работающих точек (рис. 49).
Эти явления указывают на непосредственное участие лобных долей мозга человека в активации, вызываемой речевой инструкцией; лобные доли являются частью мозговых систем, которые непосредственно вовлекаются в процессы, связанные с высшими формами активного внимания. Наличие богатых нисходящих связей лобных долей мозга с лимбической системой и ретикулярной формацией ствола является фактом, позволяющим выделить и некоторые морфофизиологические основы участия лобных долей в этих высших формах активации.
При психофизиологических исследованиях больных с поражением лобных долей мозга многолетними и тщательными исследованиями Е. Д.Хомской и ее сотрудников был установлен ряд фундаментальных фактов (Е.Д.Хомская, 1960, 1965, 1966, 1969, 1971, 1972; Е.Д.Хомская, А.Р.Лурия, 1970; Е.Д.Артемьева, 1965, 1966; О.П.Барановская, 1966, 1968; Э.Г.Симерницкая, 1966, 1970; и др.).
Как уже упоминалось, массивные поражения верхних отделов ствола среднего диэнцефального мозга и лимбической области могут привести к нарушениям первичных основ внимания — ориентировочного рефлекса, который либо угнетается и легко угасает, либо, напротив, не подчиняется факторам привыкания. Характерно, однако, что в этих случаях привлечение внимания к сигналу путем речевой инструкции может в известных пределах компенсировать этот дефект и усиливать и стабилизировать электрофизиологические и вегетативные компоненты ориентировочного рефлекса. Аналогичное можно наблюдать и у больных с поражениями задних отделов мозга, у которых патологическое состояние коры может делать физиологическую основу внимания неустойчивой, но у которых, однако, речевая инструкция, данная больному, сразу же повышает тонус коры и в определенной мере компенсирует дефект. Настойчивую работу над компенсацией дефекта можно наблюдать у этих больных и в процессе восстановительного обучения, она, безусловно, требует напряженности и стойкости произвольного внимания.
Совершенно иное можно наблюдать у больных с массивными поражениями лобных долей мозга.
Как уже говорилось, элементарные ориентировочные рефлексы у этих больных часто оказываются патологически усиленными, и именно это мешает им целенаправленно выполнять задания.
Другая картина обнаруживается у этих больных, если наблюдать их реакции на последовательно предъявляемые сигналы. Значительная часть больных этой группы обнаруживает в этих случаях большую нестойкость внимания, и уже через несколько предъявлений одних и тех же раздражителей вегетативные и электрофизиологические компоненты ориентировочного рефлекса исчезают.
Отличие больных с поражениями лобных долей от больных с очаговыми поражениями в задних отделах мозга заключается, однако, в другом: если у последних больных привлечение произвольного внимания дополнительной инструкцией (например, инструкцией «считать сигналы», «следить за их изменением» и т.п.) сразу же восстанавливает и стабилизирует угасшие компоненты ориентировочного рефлекса, то у больных с поражением лобных долей мозга этого не происходит. Инструкция, которая должна повысить активность больного, не приводит ни к каким сколько-нибудь устойчивым сдвигам в системе ориентировочных реакций.
Это проявляется в целой серии показателей процессов активации.
Так, если в норме многократное повторение сигнала приводит к угашению сосудистой ориентировочной реакции, достаточно предложить испытуемому следить за изменениями интенсивности или длительности сигнала или считать количество сигналов, сосудистые реакции, являющиеся компонентом ориентировочного рефлекса, сразу же восстанавливаются и остаются устойчивыми в течение длительного времени. Этого нет у больных с массивными поражениями лобных долей мозга; сосудистые реакции на повторяющиеся сигналы у них быстро угасают, и речевая инструкция, в норме мобилизующая внимание, не приводит у них ни к какому эффекту: кривая плестизмограммы остается неизменной.
Далее, в норме первые предъявления сигналов вызывают отчетливые сдвиги в электроэнцефалограмме в виде депрессии альфа-ритма и повышения высоких частот спектра; многократное повторение сигналов приводит к явлениям привыкания, т.е. к исчезновению этих реакций: после инструкции, привлекающей внимание к этим сигналам, ЭЭГ-компоненты ориентировочного рефлекса восстанавливаются вновь и приобретают устойчивый характер. Эта стабилизующая и активизирующая функция внимания, сохраняющаяся у больных с поражениями задних отделов больших полушарий, нарушается у больных с массивными поражениями лобных долей мозга, и если даже они и в состоянии выполнять данную им инструкцию, то это протекает без явной мобилизации внимания по ЭЭГ-показателям.
Наконец, как известно, в норме раздражение определенной модальности приводит к появлению вызванных потенциалов в соответствующих отделах мозговой коры; введение инструкции различать сигналы приводит к заметному усилению амплитуды поздних компонентов вызванных потенциалов и их большему распространению по коре головного мозга. Эти явления, подробно описанные рядом авторов, не наблюдались у больных с массивными поражениями лобных долей; хотя у них, как и у других испытуемых, сигнал вызывал отчетливый вызванный потенциал, параметры этого потенциала не менялись под влиянием инструкции, мобилизующей внимание.
Соответствующие иллюстрации приводились на рисунке 86.
Все эти данные дают полное основание предполагать, что лобные доли мозга принимают ближайшее участие в повышении уровня бодрствования соответственно поставленным перед субъектом задачам и тем самым играют решающую роль в обеспечении высших произвольных форм внимания. Этот факт, несомненно имеющий большое значение для понимания мозговых механизмов сложных форм внимания, был детально изучен в работах Е.Д.Хомской и ее сотрудников, и сравнительные наблюдения над различными формами поражения лобных долей мозга позволили не только установить только что сформулированное положение, но и существенно уточнить его.
Оказалось, что описанное явление нарушения регуляции процессов активации посредством речевой инструкции проявлялось особенно отчетливо у больных с массивными двусторонними поражениями лобных долей мозга, захватывающими ее медиальные или медиобазальные области. Это выступало как у тех больных, которые проявляли в своем поведении уже описанные выше черты инактивности, отвлекаемости и глубокого нарушения избирательности психических процессов, так и у больных, которые клинически не проявляли заметных для внешнего наблюдения симптомов изменения поведения.
Больные с поражениями конвекситальных отделов лобных долей мозга обнаружили другую картину. После угашения ориентировочных реакций, возникавших у них под влиянием сигналов, вызвать их усиление и стабилизацию с помощью речевой инструкции оказывалось все же возможным, хотя лишь после многократного повторения инструкции. Однако в этих случаях они оказывались нестойкими и быстро исчезали.
Все эти исследования имеют большое значение для установления мозговых механизмов высших произвольных форм внимания. Они показывают, что лобные доли мозга играют важную роль в организации произвольной сознательной деятельности человека.
Глава IV ПАМЯТЬ
ПСИХОЛОГИЧЕСКОЕ СТРОЕНИЕ
Изучение механизмов памяти долгое время было малоразработанным разделом нейрофизиологии; только за последнее десятилетие появились работы в этом направлении. Мозговая организация различных форм мнестической деятельности остается в значительной степени новой главой нейрофизиологической науки.
Причиной такого положения в большой степени являются упрощенные взгляды на память, которых придерживается подавляющее большинство физиологов, и недооценка сложности структуры мнестических процессов, продемонстрированной за последние 20 — 30 лет (здесь и далее в подобных случаях иметь в виду, что первое издание книги вышло в 1973 году. — примеч. ред.) психологами.
Если резюмировать все, что было известно в психологии о природе и о материальном субстрате памяти к началу этого века, становится ясно, насколько бедна была информация, которой располагала наука.
С одной стороны, она включала положение Р.Семона и Э.Геринга о том, что память, или «способность сохранять следы», является «всеобщим свойством материи», — положение, которому нельзя отказать в правильности, но которое имеет слишком общий характер. С другой стороны, бытовали представления А. Бергсона (1896) о том, что существуют два вида памяти: «память тела» и «память духа» и что если первая относится к числу естественных явлений — тех самых, о которых говорили Семон и Геринг, — то вторая должна рассматриваться как проявление «свободной воли», способной путем духовного, волевого усилия вызывать индивидуальные следы прежнего опыта.
Сравнительно мало обогатили вопрос о природе памяти и морфофизиологические исследования первых четырех десятилетий этого века. Тщательное морфологическое исследование нервной клетки и ее связей не пошло дальше общего утверждения, что сохранение следов раз возникших возбуждений является, по-видимому, результатом наличия синаптического аппарата (Рамон-и-Кахал, 1909—1911) и что в основе его, по-видимому, лежат некоторые биохимические процессы, связанные с равновесием
ацетил-холина и холин-эстеразы, играющих основную роль в синаптической передаче импульса (де Робертис, 1964; Экклз, 1957— 1966). Работы по физиологии условного рефлекса, целиком посвященные процессам выработки и закрепления опыта, установили лишь некоторые основные физиологические факторы того, что весьма условно обозначалось как «проторение путей» или «закрепление условных связей», но фактически не сделали каких-либо существенных шагов для раскрытия природы памяти. Не достигли больших успехов и исследователи процесса научения (learning) — в основном американские бихевиористы. Несмотря на то что число публикаций, посвященных этому вопросу, исчислялось многими тысячами, вопрос о природе памяти оставался совершенно неясным. Именно это и дало основание выдающемуся американскому психологу К. С.Лешли в его известной статье «В поисках энграммы» (1950) прийти к пессимистическому заключению, что материальная природа памяти остается такой же нераскрытой, какой она была многие десятилетия назад.
Существенный сдвиг в понимании материальных основ памяти был сделан в результате работ Хидена (I960, 1962, 1964), показавшего, что сохранение следа от возбуждения связано с изменением структуры рибонуклеиновой кислоты, и проследившего закономерное возрастание содержания РНК—ДНК в ядрах, подвергавшихся интенсивному возбуждению. Работы Хидена положили начало целому потоку исследований, и положение, что молекулы РНК—ДНК являются носителями памяти, что именно они играют решающую роль как в наследственной передаче, так и в прижизненном сохранении следов памяти, вскоре стало общепризнанным.
Из этого положения был даже сделан вывод, что информация, полученная одной особью, может передаваться гуморальным путем второй особи; получившие широкую огласку (но остающиеся дискуссионными) опыты Мак-Коннелла и Джекобсона (1970) вселяли надежду на то, что материальные основы памяти будут вскоре окончательно раскрыты.
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 371; Нарушение авторских прав?; Мы поможем в написании вашей работы!