КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Ионный состав внутриклеточной и межклеточной среды вещества и мембранный потенциал
|
|
|
|
Возбудимые клетки поляризованы, т.е. по разные стороны клеточной мембраны существует разность потенциалов, называемая мембранным потенциалом клетки. Внутри клетки содержится много отрицательных зарядов, поэтому внутренняя среда клетки заряжена более отрицательно, чем наружная.
Содержание ионов в клетках позвоночного
| Ионы | Мышечные волокна (а также нейроны) позвоночного | |
| Внутриклеточная среда | Внеклеточная среда (межклеточная жидкость) | |
| Катионы | ||
| K+ | ||
| Na+ | ||
| Ca2+ | 5(в основном в связном виде) | |
| Mg2+ | ||
| Анионы | ||
| Cl- | ||
| HCO3- | ||
| (Aминокис)- |
Мембрана при этом выступает как конденсатор – две электропроводящие среды разделены изолирующим липидным слоем. Для измерения мембранного потенциала один регистрирующий электрод помещают во внеклеточную жидкость, а другой – во внутриклеточную среду, и при этом с помощью вольтметра можно зарегистрировать мембранный потенциал. Для таких исследований используют стеклянные микроэлектроды – очень тонкие стеклянные трубочки, заполненные раствором электролита. Диаметр кончика такого электрода очень мал, и его можно вводить в клетки, практически не повреждая их. Так что электрические процессы, происходящие в клетке, можно регистрировать в живой, нормально функционирующей клетке.
6. Природа и значение потенциала покоя клеток. Уравнение Нернста.(+ Na+/К+-насос)
Возбудимые клетки могут находиться в покоящемся и в возбужденном состоянии. Мембранный потенциал покоящейся клетки называется потенциалом покоя (ПП). Величина его достаточно сильно различается в разных клетках. Изменяется состояние клетки, и изменяется мембранный потенциал (и наоборот). Если внутри клетки увеличится количество положительных зарядов, то мембранный потенциал будет уменьшаться по абсолютной величине, т.е. будет происходить деполяризация (уменьшение поляризованности клетки). Если, наоборот, из покоящейся клетки удалять положительные заряды, то абсолютная величина мембранного потенциала будет увеличиваться, т.е. будет происходить гиперполяризация (увеличение поляризованности клетки). Если клетка была предварительно деполяризована, то в последнем случае будет не гиперполяризация, а реполяризация (возвращение к исходной поляризованности). Поляризовать можно любую клетку. Но для мембран возбудимых клеток (большинство нервных, мышечных и рецепторных клеток) характерно наличие порогового потенциала (порог возбудимости). Для каждого вида клеток существует определенная величина порога возбудимости. Если деполяризация клетки достигает значения порога, то развивается мощный активный ответ клетки – потенциал действия (ПД). Он возникает в результате скачкообразного изменения свойств мембраны под действием деполяризации.
Рассмотрим состояние покоя. За счет чего создается разность зарядов между внутри- и внеклеточной средой? Она создается вследствие неравномерности распределения ионов по разные стороны мембраны.!!!!!
Итак, мембрана обладает емкостными свойствами благодаря наличию электроизолирующего двойного липидного слоя. Но также мембрана обладает проводимостью, за счет того, что в липидный слой встроены специальные структуры – ионные каналы и ионные насосы, с помощью которых ионы могут проходить через мембрану и переносить с собой электрические заряды, обеспечивая таким образом возникновение и изменение мембранного потенциала (МП). Проводимость мембраны является мерой ее ионной проницаемости, т.е. чем выше проводимость, тем большее число ионов может пересечь мембрану за единицу времени под действием электрической силы – разности потенциалов.
Источником энергии для активных электрических процессов, протекающих в мембране, и элементом, обусловливающим ПП, является электрохимический потенциал. Он обусловлен двумя основными свойствами клеток:
· асимметричным распределением ионов между внутри- и внеклеточными жидкостями, которое поддерживается метаболическими процессами;
· избирательной проницаемостью ионных каналов клеточных мембран
Равновесный потенциал для того или иного иона должен увеличиваться при увеличении трансмембранного концентрационного градиента этого иона. Уравнение, связывающее отношение концентраций и мембранный потенциал, было выведено в конце XIX в. Вальтером Нернстом:
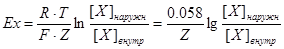 , где
, где
R – универсальная газовая постоянная;
T – абсолютная температура в градусах Кельвина;
F– число Фарадея (96500 Кл/г-экв.заряда);
Z – электровалентность иона; [X]наружн и[X]внутр – концентрация иона снаружи и внутри клетки;
Ex – равновесный потенциал для иона Х.
Электрический потенциал содержимого живых клеток (Евнутр) принято измерять по отношению к потенциалу среды (Енаружн).
Например, калиевый равновесный потенциал отрицателен, т.к. внутри К больше, чем снаружи, и логарифм величины меньше единицы отрицателен.
Обычно ПП составляет от -30 до -100 мВ. Его величина зависит от типа клетки и ионного состава окружающей среды. Если мембрана малопроницаема для какого-либо иона, то он будет оказывать меньшее влияние на величину МП. Именно относительная способность различных ионов к диффузии через мембрану определяет их вклад в МП, возникающий благодаря этой диффузии.
В мышцах, нервах и в большинстве других клеток ПП наиболее чувствителен к концентрации К+, так как проницаемость мембраны для К+ относительно велика по сравнению с остальными катионами.
Мембраны живых клеток в той или иной степени проницаемы для всех ионов, ионы будут утекать (т.н. «токи утечки»), стремясь выровнять концентрационный градиент. Для поддержания потенциала покоя и восстановления исходного состояния нейрона после разряда импульсов (так же, как и для многих других внутриклеточных процессов) необходим активный транспорт ионов через мембрану, они используют механизм активного транспорта некоторых ионов против их электрохимического градиента.
Основной механизм активного переноса ионов через мембрану представлен ферментом – Na+/К +- АТФазой, или Na+/К+-насосом.
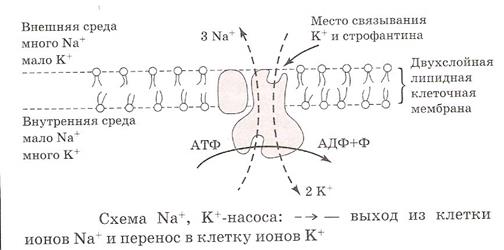 Na+/К+-насос представляет собой белковую структуру, пронизывающую насквозь мембрану клетки. Этот белок может связывать ионы Na+ с внутренней и ионы К+ – с наружной стороны мембраны. Затем он может изменять свою конформацию (структурную организацию) таким образом, что совершает как бы «кувырок» в мембране и при этом «обменивает» ионы друг на друга. На эти конформационные перестройки и на преодоление силы градиента концентраций затрачивается энергия, получаемая при расщеплении молекулы АТФ. Таким образом поддерживается электрохимический градиент для ионов натрия и калия. Этот насос является также электрогенным, т.к. удаляет из клетки больше положительных зарядов (3 иона натрия наружу), чем закачивает внутрь (2 иона калия), таким образом увеличивая поляризованность клетки. В настоящее время выявлены и изучены не только натриевые и калиевые, но также кальциевые и хлорные насосы.
Na+/К+-насос представляет собой белковую структуру, пронизывающую насквозь мембрану клетки. Этот белок может связывать ионы Na+ с внутренней и ионы К+ – с наружной стороны мембраны. Затем он может изменять свою конформацию (структурную организацию) таким образом, что совершает как бы «кувырок» в мембране и при этом «обменивает» ионы друг на друга. На эти конформационные перестройки и на преодоление силы градиента концентраций затрачивается энергия, получаемая при расщеплении молекулы АТФ. Таким образом поддерживается электрохимический градиент для ионов натрия и калия. Этот насос является также электрогенным, т.к. удаляет из клетки больше положительных зарядов (3 иона натрия наружу), чем закачивает внутрь (2 иона калия), таким образом увеличивая поляризованность клетки. В настоящее время выявлены и изучены не только натриевые и калиевые, но также кальциевые и хлорные насосы.
Мембранный потенциал создается в результате как активных, так и пассивных механизмов транспорта ионов. Степень участия тех или иных механизмов в разных клетках неодинакова, поэтому и мембранный потенциал в клетках разного типа отличается (от -40 до -90 мВ).
- Условия возникновения ПД. Закон «всё или ничего».
- Рефрактерность мембраны нейрона.
- Активный нейронный транспорт. Ионные каналы и насосы.
- Виды и значение ионных каналов в мембране возбудимых клеток.
ПД (спайк, импульс) возникает на мембранах возбудимых клеток (нервных и мышечных клеток, а также некоторых рецепторных и секреторных клеток). Возникновению ПД обычно предшествует деполяризация клеточной мембраны. Для этого необходим деполяризующий ток (как правило, это ток, создаваемый входящими положительными зарядами). Этот ток может возникать при электрическом воздействии на клетку (если, например, возбуждается соседняя тесно связанная клетка), но чаще всего при действии на клетку специальных возбуждающих веществ, о которых разговор пойдет позже.
Если величина деполяризующего тока меньше порогового значения, то возникает лишь пассивная деполяризация мембраны, нераспространяющееся возбуждение, называемое локальным ответом. Если деполяризация достигает порогового уровня, то развивается потенциал действия. По достижении порогового потенциала дальнейшая деполяризация нарастает лавинообразно, потенциал становится все менее отрицательным и в какой-то момент даже положительным (т.н. овершут).
В норме можно зарегистрировать либо максимальный электрический ответ мембраны (ПД), либо очень небольшой локальный ответ, но не какие-нибудь промежуточные по величине реакции. Поэтому говорят, что ПД подчиняется закону «все или ничего», т.е. амплитуда ПД всегда максимальна и не зависит от силы раздражителя.
Еще одна характерная особенность ПД – быстрая реполяризация до уровня покоя после достижения максимума. Длительность ПД может быть разной: в некоторых нервных волокнах она составляет миллисекунды, а в сердечной мышце – около 0,5 сек. При уменьшении интервала между двумя ПД величина второго из них становится все меньше и меньше. Если повторный стимул (деполяризующий ток) подается очень быстро после окончания первого ПД, то второй ПД не развивается. Этот период состояния нейрона называется периодом рефрактерности.
В 1949 г. А. Ходжкин и Б. Кац выдвинули т.н. натриевую гипотезу природы ПД, согласно которой фаза подъема и овершут ПД обусловлены временным повышением проницаемости мембраны для Na+ и входом этого иона в клетку.
В пользу натриевой гипотезы говорит следующее:
1. Содержание Na+ во внеклеточной среде примерно в 10 раз больше, чем в клетке, поэтому ЕNa составляет 50–60мВ. Направление действующей на ионы Na+ электродвижущей силы (ЭДС = Ем - ЕNa) таково, что эти ионы стремятся пройти в клетку.
2. Поскольку ионы Na+ заряжены положительно, то их вход в клетку должен привести к деполяризации, что и происходит.
3. На высоте овершута ПД приближается к равновесному Na+-потенциалу. Он составляет 58 мВ.
4. Величина овершута зависит от содержания ионов Na во внеклеточной среде, причем эта зависимость соответствует теоретической, которую можно рассчитать из уравнения Нернста для ЕNa.
Показано, что при действии деполяризующего тока на клетку наблюдается сначала очень быстрый входящий ток, а спустя 1–2 мсек он сменяется более медленно развивающимся выходящим током. Первый определяется входящими ионами Na+ и отвечает за быструю деполяризацию, второй определяется выходящими ионами К+ и отвечает за реполяризацию.
Входящий натриевый ток. При возбуждении мембраны ионы Na+ входят через специальные каналы в мембране, избирательно проницаемые для этих ионов. Na+-каналы активируются (т.е. открываются и пропускают ионы) в ответ на деполяризацию, при этом они избирательно пропускают преимущественно ионы Li+ и Na+. Такое свойство обусловлено особой молекулярной структурой каналов, благодаря которой они играют роль селективных фильтров. Белковые молекулы, формирующие каналы, могут изменять свою конформацию (структурную организацию) под действием изменений МП, в результате чего каналы открываются (активируются), закрываются и инактивируются. Предполагается воротный механизм работы Na+-канала (рис 2.8). Блокатором этих каналов (веществом, связывающимся с молекулами каналов и блокирующим их работу), является тетродотоксин, яд, выделенный из внутренних органов рыбы иглобрюха. Он как бы затыкает канал. В лабораторных экспериментах его используют для блокады передачи нервных импульсов.
Цикл Ходжкина. При деполяризации число открытых Na+-каналов увеличивается. Увеличение Na+-проводимости лежит в основе тех самоусиливающихся процессов, из-за которых ПД подчиняется закону «все или ничего». При действии деполяризующего стимула сначала происходит открывание лишь небольшого числа Na+-каналов. Через них некоторое количество ионов Na+ войдет в клетку. Они принесут положительные заряды и дополнительно деполяризуют клетку. Это приведет к открыванию дополнительных Na+-каналов и т.д. Такой замкнутый круг, связывающий МП и Na+-проводимость, называется циклом Ходжкина (пример положительной обратной связи).
Выходящий калиевый ток отвечает за реполяризацию клетки. По мере того, как МП становится все более положительным, увеличивается электродвижущая сила (ЭДС), действующая на ионы К+, т.е. ионы К+ стремятся покинуть клетку. Каналы, отвечающие за этот ток, также активируются при деполяризации. Но они открываются медленнее, чем Na+-каналы, поэтому ПД успевает развиться без помех. Отличие Na+- и К+- каналов в том, что при деполяризации Na+-каналы быстро открываются и быстро инактивируются под действием деполяризации, далее Na+-каналы могут вновь открыться только когда мембрана будет опять реполяризована до исходного уровня МП. А К+-каналы открываются медленно и не инактивируются при деполяризации, а закрываются только тогда, когда мембрана реполяризуется до исходных значений.
Отсюда возникает такое состояние возбудимой клетки, как рефрактерность. Во время ПД происходит два процесса, в результате которых нейрон не способен какое-то время генерировать следующий потенциал действия. Во-первых, инактивация натриевых каналов достигает наибольшего уровня во время фазы спада ПД, и на ее устранение необходимо несколько миллисекунд. Во-вторых, благодаря активации калиевых каналов, проницаемость для этих ионов очень высока на спаде ПД и постепенно возвращается к уровню покоя. При таких условиях для деполяризации потребовалось бы очень большое увеличение проницаемости для ионов натрия.
Оба перечисленных фактора приводят мембрану клетки в состояние абсолютной рефрактерности – полной невозбудимости, когда даже сверхпороговые стимулы не могут вызвать повторного ответа в виде ПД. После абсолютной рефрактерности наступает период относительной рефрактерности, в ходе которого порог постепенно снижается до своего нормального уровня, натриевые каналы восстанавливаются из инактивированного состояния, а калиевые каналы закрываются. Во время относительной рефрактерности деполяризующий сигнал достаточно большой амплитуды может активировать такое число Na+-каналов, чтобы, несмотря на большое число еще открытых К+-каналов, возник ПД. Однако вследствие высокой К+-проводимости и значительной остаточной Na+-инактивации МП в точке максимума не будет столь близок к ЕNa, а следовательно, амплитуда ПД будет меньше.
Важно помнить, что активный транспорт ионов через мембрану (напр., Na+/К+-насос) не отвечает непосредственно за фазы деполяризации и реполяризации ПД, но необходим для поддержания ионных градиентов, которые ответственны за возникновение мембранных токов.
Существуют также и другие электровозбудимые каналы. Во многих возбудимых клетках есть потенциалзависимые Са++-каналы. Однако Са++-токи не вносят существенного вклада в развитие ПД. Са++ в клетке, как правило, выступает в качестве посредника, запускающего многие внутриклеточные процессы, секрецию и т.д.
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 5604; Нарушение авторских прав?; Мы поможем в написании вашей работы!