КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Позиционные сигналы и пролиферация клеток
|
|
|
|
НЕКОТОРЫЕ СТИМУЛИРУЮЩИЕ И ИНГИБИРУЮЩИЕ ПУТИ ВЛИЯНИЯ НА ПРОЛИФЕРАЦИЮ КЛЕТОК С ПОМОЩЬЮ АКТИВАЦИИ ИЛИ ИНАКТИВАЦИИ ФОСФОРИЛИРОВАНИЯ рRB
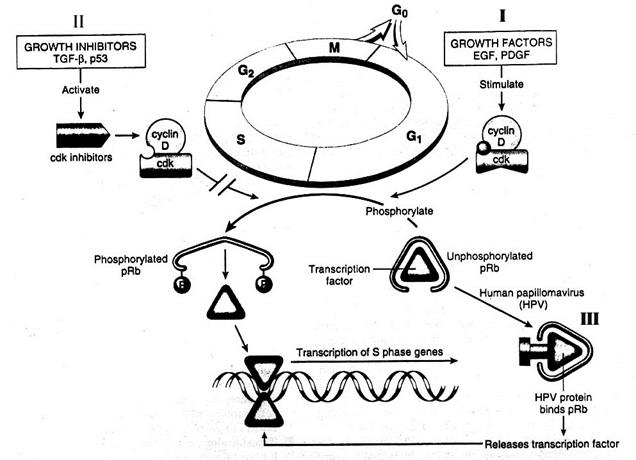 |
Ростстимулирующие и ростингибирующие факторы. Механизм действия на клеточный цикл (по V. Cumar, R. S. Cotran, S. L Robbins, 1997)
· Стимулируя или блокируя процесс фосфорилирования pRb, можно, соответственно, активировать или подавлять клеточный цикл
· Факторы роста: эпидермальный (EGF), тромбоцитарный (PDGF) являются таковыми именно потому, что стимулируя активность сdk, усиливают процесс фосфорилирования pRb и вхождение клетки в S-фазу (I)
· Ингибиторы роста: трансформирующий ростовый факторβ (TGFβ), белок р53 индуцируют синтез протеинов - cdk inhibitors (р27, р16, р21), которые, связываясь с комплексом сdk - циклин, инактивируют сdk, блокируя тем самым клеточное деление (II)
· Некоторые ДНК-содержащие вирусы, например, вирус папилломы человека (HPV) несут протеины, которые альтернативно E2F cвязываются с нефосфорилированным pRb, исключая для него возможность образовывать комплексы и секвестировать E2F. В результате E2F перманентно пребывает в несвязанном состоянии, поддерживая митотический цикл (III).
· Размножение нормальных клеток помимо факторов роста регулируется и позиционными сигналами, опосредуемыми контактами клеток друг с другом и контактами клеток с внеклеточным матриксом. Поскольку такие контакты осуществляются при участии молекул адгезии, соответственно, кадгеринов и интегринов, последним принадлежит важная роль в регуляции клеточной пролиферации (Ю.А.Ровенский,1998; Е.Д. Свердлов, 2001).
· Подобная регуляция осуществляется при участии цитоплазматических белков - катенинов (плакоглобинов), обеспечивающих связь кадгеринов с активными микрофиламентами клетки, содержащими молекулы винкулина и актина.
· Наиболее изучен регуляторный механизм на примере E-кадгерин-катенинового комплекса. Посттрансляционная регуляция этого комплекса реализуется благодаря фосфорилированию β-катенина по тирозину. Присоединяясь к цитоплазматическому домену кадгеринов, β-катенин подвергается воздействию подмембранной тирозиновой протеинкиназы (ТК; рр60src), активируется и участвует в интеграции с цитоскелетными структурами контактирующих клеток.
· Чем сильнее выражена экспрессия кадгеринов на клеточной мембране, тем больше связывается с их цитоплазматическим доменом молекул β-катенина, тем больше этих молекул подвергается фосфорилированию под действием протеинкиназы.
· За фосфорилирование β-катенина с подмемебранными тирозинкиназами (ТК) конкурирует GSK-3 (киназа гликогенсинтазы 3 β), которая также переводит β-катенин в фосфорилированную форму.
· Фосфорилированный GSK-3 β-катенин отличается нестабильностью и приобретает чувствительность к белку-супрессору АРС. Под влиянием этого белка он подвергается деградации.
· Таким образом, в покоящейся клетке содержание свободного (нефосфорилированного) β-катенина лимитировано: а) его связыванием с кадгерином с последующим фосфорилированием подмембранной тирозинкиназой; б) его фосфорилированием под влиянием GSK с последующей АРС-опосредованной деградацией.
· Это лимитирование может быть нарушено: а) при снижении экспрессии на клеточной поверхности кадгерина; б) при блокаде активности GSK-3; в) при мутации гена супрессорного белка АРС, делающей его неспособным к осуществлению деградации фосфорилированного β-катенина
· В любом случае накопление в клетке нефосфорилированного β-катенина приводит к важным регуляторным сдвигам в отношении клеточного роста, инвазивности и направленной миграции клеток.
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 428; Нарушение авторских прав?; Мы поможем в написании вашей работы!