КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Сосудистая ткань 1 страница
|
|
|
|
МЫШЦА
6,,
I
МЯГКИЕ ТКАНИ
Ткани можно разделить на две категории: жесткие и мягкие. К первым относятся кости, а также зубы, ногти и волосы. К мягким тканям относятся сухожилия, связки, мышцы, кожа и большинство других тканей (Mathews, Stacy и Hoover, 1964). Мягкие ткани разделяют на две группы: сократительные и несократительные.
Свойства мягких тканей. Мягкие ткани отличаются своими физическими и механическими характеристиками (рис. 5.7). Как сократительные, так и несократительные ткани являются растяжимыми и эластич-
Наука о гибкости
 30 г ными, однако первые являются
30 г ными, однако первые являются
| А Фасция В Сухожилие С Склера D Роговица Е Кожа F Аорта G Задняя часть связки |
еще и сжимаемыми. Сокращаемость представляет собой способность мышцы укорачиваться и производить напряжение вдоль своей длины. Растяжимость — это способность мышечной ткани растягиваться в ответ на приложенную извне силу. Чем меньше силы, производимые в мышце, тем больше степень растяжения.
| 30 40 50 Нагрузка, % |
| Рис. 5.7. Кривые нагрузки-деформации для различных соединительных тканей (Soden, Kershaw, 1974) |
Взаимосвязь между механическими свойствами мягких тканей и растягиванием. Чем выше жесткость мягкой ткани, тем большую силу следует приложить, чтобы вызвать ее удлинение. Ткань, имеющая малую степень жесткости, не способна противостоять растягивающему усилию в той же мере, что и ткань с высокой степенью жесткости, и поэтому для производства такой же деформации требуется значительно меньшая сила, а мягкие ткани с более высокой степенью жесткости менее подвержены травмам (включая разрывы связочной ткани и сократительной, или мышцы).
Мягкие ткани не являются совершенно эластичными. Если превышен предел эластичности, то после прекращения действия силы они не способны восстановить свою исходную длину. Разница между исходной и новой длиной называется количеством потерянной эластичности. Эта разница коррелирует с минимальным повреждением ткани. Следовательно, в случае незначительного растяжения мягкие ткани не восстанавливают исходную длину после устранения чрезмерной нагрузки, что ведет к постоянной нестабильности сустава.
Возникает естественный вопрос: надо ли для развития гибкости растягиваться до предела эластичности или следует только слегка превышать его? Большинство авторитетов рекомендуют растягиваться до появления чувства дискомфорта или напряжения, но не боли. Однако в чем заключается разница между дискомфортом и болью? Значение этих понятий в медицине (и других дисциплинах) можно интерпретировать по-разному, в зависимости от того, кто осуществляет интерпретацию (de Jong, 1980). В 1979 г. была создана Международная ассоциация по изучению боли с целью разработать общеприемлемое определения понятия «боль», а также систему классификации болевых синдромов. Было дано определение боли и названы еще 18 общих терминов (de Jong, 1980, Merskey, 1979). Нас интересуют только три:
 Глава 5 ■ Механические и динамические свойства мягких тканей
Глава 5 ■ Механические и динамические свойства мягких тканей
Боль — неприятные ощущения, связанные с действительным или возможным повреждением ткани или охарактеризованные как подобное повреждение.
Болевой порог — наименьшая интенсивность стимула, при которой человек испытывает боль.
Уровень болевой толерантности — наибольшая интенсивность стимула, вызывающая боль, которую готов перенести человек.
Исходя из этих определений, большинство специалистов делают вывод, что растягиваться следует по меньшей мере до болевого порога. Но так как эти три определения основаны на субъективных факторах, тренеры не могут установить уровень болевого порога у своих подопечных. Такого понятия, как «средний человек», не существует, каждый человек уникален в своих ощущениях и восприятиях, которые к тому же постоянно изменяются.
Особое внимание необходимо обратить на следующее. У лиц, проходящих реабилитацию и восстанавливающих поврежденные ткани, еще до возникновения боли может быть достигнуто состояние, при котором возможен разрыв этих тканей. Поэтому при воздействии на них следует соблюдать особую осторожность.
Кроме того, возникает еще один вопрос: находится ли точка дискомфорта ниже, на уровне или выше предела эластичности? Согласно результатам исследований вид силы, ее продолжительность, а также температура ткани во время и после растягивания определяют, является ли удлинение постоянным и обратимым.
Соотношение длина-напряжение и нагрузка-деформация. Длина мягкой ткани зависит от отношения внутренней силы, развиваемой тканью, ко внешней силе, обусловленной сопротивлением развитию внутренней силы или нагрузкой. Если внутренняя сила превышает внешнюю, ткань сокращается. Если же внешняя сила превышает внутреннюю, ткань удлиняется.
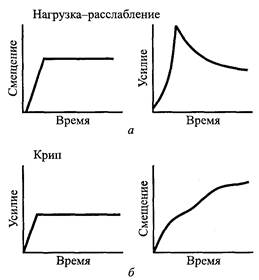
Нагрузка-расслабление и крип при пассивном напряжении. Живые ткани характеризуются наличием зависимых от времени механических свойств. К ним относятся нагрузка-расслабление и крип. Если находящуюся в состоянии покоя мышцу внезапно растянуть и постоянно удерживать достигнутую длину, то через некоторое время произойдет медленное снижение напряжения. Это поведение называют нагрузка-расслабление (рис. 5.8, а). С другой стороны, удлинение, которое происходит при воздействии постоянной силы или нагрузки, называется крип (рис. 5.8, б).
Каким же образом эти зависимые от времени механические свойства действуют на мышечные клетки и соединительные ткани. Несомненный интерес представляют следующие вопросы:
• Как передается растягивающая сила через саркомер и структуры различных соединительных тканей?
• Как влияет растягивающая сила на сарколемму, саркоплазму и цитос-келет саркомера?
• Где и через какие структуры саркомера осуществляется явление крип и нагрузка-расслабление?
Наука о гибкости
|
• Каково взаимоотношение (если таковое существует) между крипом и нагрузкой-расслаблением в саркомере и градиентами давления, потоком жидкости и потенциалами течения структур различных соединительных тканей?
| Рис.5.8. Реакция тканей на усилие: нагрузка-расслабление имеет место при снижении усилия в случае, если длина ткани сохраняется постоянной (а); крип представляет собой удлинение, имеющее место при действии постоянной силы в течение периода времени (б) |
Молекулярный механизм эластичной реакции соединительной ткани. Соединительные ткани представляют собой сложные материалы, которые, соединяясь, образуют длинные гибкие цепочки. Двумя важнейшими переменными, влияющими на жесткость (или эластичность) соединительных тканей, являются расстояние между поперечными соединениями и температура. Представим, например, длинную гибкую молекулу, состоящую из определенного числа сегментов. Количество сегментов обозначим буквой п. Каждый сегмент имеет определенную длину, обозначенную буквой а. Допустим, что каждый сегмент является жестким, тогда как суставы между сегментами — гибкими. Также допустим, что молекулы сегментов свободно передвигаются.
Все молекулы двигаются сравнительно хаотично. Однако при снижении температуры их движение становится не таким свободным. При достижении температуры абсолютного нуля (-273° С) движение прекращается. Вследствие хаотичного движения молекул в определенный момент расстояние от одного конца сегмента до другого может иметь значение от О (если концы соприкасаются) до па (если молекулы вытянуты). Наиболее вероятная длина молекулы равна п1/2а.
В «нормальном» состоянии молекулярные цепочки сети продолжают двигаться. Расстояние между концами конкретной цепочки изменяется, однако среднее расстояние в образце, содержащем много цепочек, всегда будет п1/2а.
Рассмотрим рис. 5.9. Допустим, что на соединительную ткань действует внешняя растягивающая сила (5.9, а). Сетка подвергнется деформации (рис. 5.9, б), и цепочки расположатся в направлении растяжения. Следовательно, цепочки, расположенные в направлении растягивающей силы (например, АВ), будут иметь среднюю длину больше п"2а. Цепочки же, расположенные поперек направления растяжения (ВС), будут иметь среднюю длину меньше п"2а. В результате этого расположение уже не является хаотичным. После устранения действия силы цепочки снова принима-
| |  |
| Глава 5 ■ Механические и динамические свойства мягких тканей |
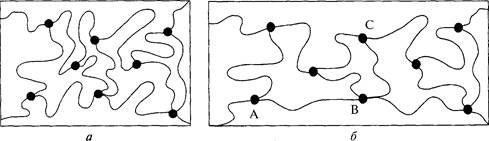
Рис. 5.9. Диаграмма резинового полимера. Синусоидой показаны молекулы полимера, точками — поперечные соединения (Alexander, 1988)
ют хаотичную конфигурацию. Таким образом, соединительная ткань восстанавливает свою исходную форму; она эластично возвращается к исходному уровню.
Р.М.Александер (1988) пишет:
«Теория, созданная на основании этих идей, позволяет определить величину силы, необходимую для уравновешивания деформированной сети и, следовательно, модуля упругости. Модуль сдвига G и модуль Юнга Е можно получить из уравнения
G = NkT = E/3,
где N — количество цепочек на единицу объема материала; к — постоянная Больцмана; Т — абсолютная температура. Особую роль играет число цепочек. Если имеется большее число поперечных соединений, разделяющих молекулы на множество более коротких цепочек, жесткость материала увеличивается. Кроме того, модуль пропорционален абсолютной температуре, поскольку энергия, связанная со скручиванием (сплетением) молекул, увеличивается при повышении температуры. Также по мере повышения температуры увеличивается давление газа при постоянном объеме, так как при этом увеличивается количество кинетической энергии молекул».
Данные исследований, касающиеся растягивания соединительной ткани. При воздействии растягивающей силы на соединительную ткань или мышцу ее длина увеличивается, а площадь поперечного сечения (ширина) уменьшается. Существуют ли такие виды сил или состояний, при которых прилагаемая сила может обеспечить оптимальное изменение соединительной ткани? Сапега с коллегами (1981) отмечают следующее:
«При непрерывном воздействии растягивающих сил на модель организованной соединительной ткани (сухожилие) время, в течение которого происходит необходимое растягивание ткани, обратно пропорционально прилагаемым силам (C.G.Warren,
Наука о гибкости
Lehmann, Koblanski, 1971,1976). Таким образом, при использовании метода растягивания с небольшой силой требуется больше времени, чтобы достичь такой же степени удлинения, как при использовании метода растягивания с большой силой. Однако процент удлинения ткани, который имеет место после устранения растягивающего усилия, оказывается выше при использовании продолжительного метода с небольшой силой (C.G.Warren и др., 1971, 1976). Кратковременное растягивание с большой силой способствует восстанавливающейся деформации эластичной ткани, тогда как продолжительное растягивание с небольшой силой —; остаточной, пластической деформации (С. G. Warren и др., 1971, 1976; Labon, 1962). Результаты лабораторных исследований показывают, что при постоянном удлинении структур соединительной ткани имеет место определенное механическое ослабление, хотя разрыв и не происходит (C.G.Warren и др., 1971, 1976). Степень ослабления зависит от способа растягивания ткани, а также от степени растяжения.
Температура существенно влияет на механическое поведение соединительной ткани в условиях растягивающего напряжения. При повышении температуры ткани степень жесткости уменьшается, а степень растяжимости увеличивается (Laban, 1962; Rigby, 1964). Если температура сухожилия превышает 103°Ф, количество постоянного удлинения в результате данного количества исходного растягивания увеличивается (Laban, 1962; Lehmann, Masock, Warren u Koblanski, 1970). При температуре около 104° Ф происходит термальное изменение микроструктуры коллагена, которое значительно усиливает расслабление вязкости после нагрузки коллагеновои ткани, что обеспечивает более высокую пластическую деформацию при растягивании (Mason и Rigby, 1963). Механизм, лежащий в основе этого термального изменения, пока не известен, однако предполагают, что происходит частичная дестабилизация межмолекулярной связи, усиливающая вязкие свойства текучести коллагеновои ткани (Rigby, 1964).
Если соединительную ткань растягивают при повышенной температуре, условия, в которых ткань может охладиться, в значительной мере могут повлиять на качество удлинения, которое остается после устранения действия растягивающего напряжения. После растяжения разогретой ткани сохраняющаяся растягивающая сила во время охлаждения ткани значительно увеличивает относительную пропорцию пластической деформации по сравнению с разгрузкой ткани при все еще повышенной температуре (Lehmann и др., 1970). Охлаждение ткани до устранения напряжения позволяет коллагеновои микроструктуре больше рестабилизироваться к ее новой длине (Lehmann и др., 1970).
Глава 5- Механические и динамические свойства мягких тканей
При растягивании соединительной ткани при температурах, находящихся в обычных терапевтических пределах (102-110° Ф), количество структурного ослабления, обусловленное данным количеством удлинения ткани, обратно пропорционально температуре (C.G.Warren и др., 1971, 1976). Это явно связано с прогрессивным увеличением свойств вязкого течения коллагена по мере увеличения температуры. Вполне возможно, что термальная дестабилизация межмолекулярной связи обеспечивает удлинение при меньшем структурном повреждении.
Факторы, влияющие на упруговязкое поведение соединительной ткани, можно обобщить, отметив, что эластичной, или обратимой, деформации больше всего способствует кратковременное растягивание с большой силой при нормальной или несколько пониженной температуре тканей, тогда как пластическому, или постоянному, удлинению в большей степени способствует более длительное растягивание с меньшей силой при повышенной температуре, если только охлаждение ткани осуществляется до устранения напряжения. Кроме того, структурное ослабление, обусловленное остаточной деформацией ткани, оказывается минимальным, когда продолжительное воздействие небольшой силы сочетают с высокими температурами, и максимальным — при использовании больших сил и более низких температур. Эти данные обобщены в табл. 5.1-5.3».
Исследования, проводившиеся другими учеными (Becker, 1979; Glarer, 1980; Light и др., 1984), также показывают, что растягивание при низких-средних уровнях напряжения, является действительно эффективным.
Таблица 5.1. Факторы, влияющие на пропорцию пластичного и эластичного растяжения
Фактор
Эластичное растяжение
Пластичное растяжение
Количество прикладываемого усилия Высокое усилие Невысокое усилие
Продолжительность прикладываемого Небольшая Большая
усилия
Температура ткани Низкая Высокая
Таблица 5.2. Факторы, влияющие на упруговязкое поведение соединительной ткани
Эластичная деформация
Вязкая (пластическая) деформация
Кратковременное растягивание Продолжительное растягивание
с большим усилием с небольшим усилием
Нормальная или несколько Повышенная температура, охлаждение
пониженная температура тканей осуществляется до устранения напряжения

Мы достаточно много знаем о различных механических свойствах мышц, так как их интенсивно исследовали многие ученые на протяжении не одного десятилетия. Существует множество причин, ввиду которых необходимо изучать механические свойства мышцы. Прежде всего, таким образом мы можем лучше понять как механические реакции всей мышцы, так и механические свойства сократительных компонентов (Zierler, 1974). В данном контексте необходимость изучения механических свойств мышцы объясняется тем, что таким образом мы можем понять и определить факторы, ограничивающие гибкость, а также лучшие способы ее увеличения.
Соединительные ткани мышц. Мышца состоит из трех независимых компонентов или элементов, которые можно классифицировать либо как эластичные, либо как вязкие. Важность этих компонентов объясняется тем, что они противодействуют деформации и, таким образом, играют важную роль в определении уровня гибкости человека. Эластичные компоненты в ответ на изменение длины производят возвращающую силу. Вязкие компоненты производят усилие в ответ на интенсивность (скорость) и продолжительность изменения длины. Тремя механическими компонентами являются:
1. Параллельный эластичный компонент (ПЭК).
2. Серийный эластичный компонент (СЭК).
3. Сократительный компонент (СК).
Параллельный эластичный компонент
Компонент, отвечающий за пассивное растягивающее напряжение в мышце, называется параллельным эластичным компонентом (ПЭК). Его название объясняется тем, что он располагается параллельно сократительному механизму (рис. 5.10). Если мышцу удалить из тела, то ее длина сок-
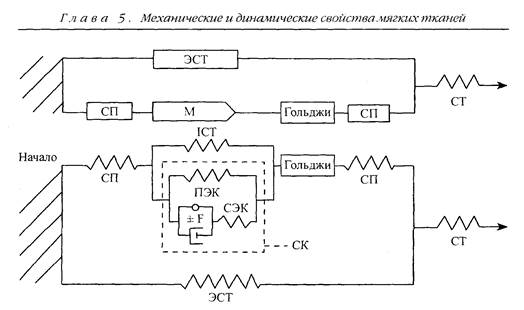
Рис. 5.10. Диаграмма упруговязких компонентов мышечного волокна и окружающей соединительной ткани с нервно-сухожильным веретеном. СП — сухожильные пучки; ЭСТ — экстрасоединительная ткань; М — мышечное волокно; Гольджи — нервно-сухожильное веретено; ПЭК, СК, СЭК, ICT — внутримышечная соединительная ткань; СТ — сухожилие
ратится примерно на 10 % по сравнению с первоначальной (Garamvolgyi, 1971). Такое сокращение длины мышцы не зависит от пассивного сокращения. Длина изолированной, несокращенной мышцы называется ее длиной равновесия; это укорачивание означает, что мышцы находятся под напряжением при нетронутой длине. Длина in situ несокращенной или нерастянутой мышцы называется длиной покоя и обозначается R1 или L0.
Находящаяся в покое мышца эластичная и противодействует удлинению. При длине, уступающей длине равновесия (0,90 L0), напряжение покоя отсутствует и ПЭК слабый. Однако когда нестимулированную мышцу растягивают, она развивает напряжение нелинейным образом. Другими словами, при начальном растягивании производится невысокое напряжение, которое по мере продолжения растягивания значительно увеличивается.
Первоначально считали, что ПЭК состоит, главным образом, из сарколеммы, саркоплазмы и эластичных волокон — эпимизия, перимизия и эн-домизия. Спустя некоторое время Х.Е. Хаксли и Хенсон (1954) выдвинули предположение об S-филаменте, который, как считали, соединял концы ак-тиновых филаментов на обоих концах. Однако уже через год Х.Е. Хаксли (1957) исключил из своей модели мышцы S-филаменты без каких-либо объяснений. Другая трактовка пассивного напряжения покоя была связана с электростатической силой. Например, известно, что объем мышечных волокон остается постоянным даже при растягивании мышцы. Площадь же поперечного сечения (ширина) мышцы должна уменьшиться, как и расстояние между филаметами актина и миозина, по мере их приближения друг к другу. Однако если между филаментами существует электростатическая сила взаимного отталкивания, то, чтобы сблизить филаменты, дол-
 Наука о гибкости
Наука о гибкости
жна быть выполнена работа. Следовательно, должна существовать сила, способствующая сохранению расположения филаментов. Таким образом, сила, необходимая для сближения филаментов, преодолевающая силу взаимного отталкивания, будет представлена напряжением покоя или «параллельным» сопротивлением растяжению (Davson, 1970; Н.Е. Huxley, 1967). Хотя электростатическая сила может способствовать напряжению покоя при высокой степени растягивания, исследования показывают, что она не может быть доминирующим источником этого процесса.
Как мы уже знаем из главы 3, основным источником эластичности мышц является титин. Это доказательство было получено в результате разрушения филаментов титина во время регистрации степени напряжения мышцы под нагрузкой. В первом исследовании титин преимущественно разрушили радиацией (Horowitz и др., 1986). Результатом оказалось снижение напряжения покоя. Год спустя Горовитц и Подольски (19876) опубликовали данные другого исследования, подтверждающие гипотезу, что эластичные филаменты титина обеспечивают большую часть напряжения покоя в мышце. В другом исследовании (Yoshioka и др., 1986) титин преимущественно разрушали путем контролируемого протеолиза, использования гидролитических (пищеварительных) ферментов. Снова отмечали снижение напряжения покоя. Еще в одном, более позднем исследовании (Funatsu, Higuchi, Ishiwata, 1990) также наблюдали снижение напряжения покоя при деградации титина в результате ферментного сжигания (гезолином плазмы).
Что же происходит, когда саркомер растягивается и отпускается? Как было показано в главе 3, длина актинового тонкого и толстого (миозинового) филаментов во время растягивания не изменяется. Они просто скользят относительно друг друга (теория скольжения филаментов). При растягивании саркомер противодействует силе деформации напряжением покоя. Вначале напряжение покоя среднее. После значительного растягивания оно резко увеличивается и противодействует последующему растяжению (эластичная жесткость). Ответственным за такое поведение является титин. После прекращения растягивания растянутые филаменты титина возвращаются в исходное положение. Таким образом, титин может накапливать потенциальную энергию.
Если титин способствует напряжению покоя, то что можно сказать о небулине? Исследования показывают, что небулин представляет собой набор нерастяжимых филаментов, прикрепленных к одному концу Z-линии, и что его филаметы идут параллельно филаментам титина (Wang, Wright, 1988). Таким образом, небулин может быть потенциальным четвертым фи-ламентом саркомера. Согласно исследованиям, при расщеплении небулина напряжение покоя не снижается (Funatsu, Higuchi, Ishiwata, 1990). Следовательно, небулин не обеспечивает эластичность.
Серийный эластичный компонент
При растягивании мышцы сократительный компонент (т. е. филаменты актина и миозина и их поперечные мостики), ПЭК и СЭК способствуют развитию напряжения. Серийный эластичный компонент (СЭК) полу-
| |
|
| Глава 5 • Механические и динамические свойства мягких тканей |
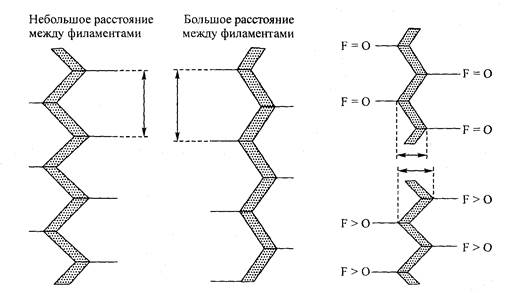
Рис. 5.11. Влияние изменения расстояния между филаментами на структуру Z-линии. Z-линия осуществляет эти изменения, меняя угол между соприкасающимися элементами
Рис. 5.12. Сила, действующая на тонкие филаменты (F), усиливает зигзагообразный паттерн структуры Z-линии, что приводит к изменению эффективной «ширины» последней. Подобные напоминающие гармошку изменения могут обусловить серийную эластичность
чил такое название потому, что эластичные компоненты располагаются непосредственно в соответствии с сократительными компонентами (см. рис. 5.10). СЭК выполняет важную функцию сглаживания резких изменений мышечного напряжения. Одной из главных анатомических частей, образующих СЭК, по мнению специалистов, является сухожилие. Следует отметить, что, по мнению Поллака (1990), Z-линия также может представлять собой определенный источник серийной эластичности саркомера. Эта функция осуществляется тонкими филаментами, которые тянут Z-линию. Например, когда сила, действующая на тонкие филаменты, передается на Z-линию, степень латерального разделения между филаментами снижается (рис. 5.11). Вместе с тем чтобы адаптировать сокращение межфила-ментного пространства, угол изгиба в структуре Z-линии становится более острым (рис. 5.12). Следовательно, Z-линия эффективно утолщается, создавая своеобразную «эластичность».
Сократительный компонент
Способность мышцы увеличивать напряжение называется сократительным компонентом (СК). СК мышцы можно рассматривать как генератор напряжения. Он состоит из филаментов и их поперечных мостиков. Если напряжение пропорционально числу химических связей, установленных между двумя филаментами, то с увеличением степени взаимного перекрытия филаментов большее число участков связывания может взаимо-
Наука о гибкости
действовать и большее напряжение может быть развито. Предполагают, что максимальное сократительное напряжение развивается при длине сар-комера, обеспечивающей наиболее полное отдельное перекрытие фила-ментов актина и миозина. При большей длине мышцы число поперечных соединений уменьшается по мере снижения степени перекрытия филамен-тов, что приводит к снижению напряжения. Если продолжать растягивание, развиваемое напряжение может уменьшаться до тех пор, пока оно не превысит напряжение, наблюдаемое в пассивной мышце. Это происходит потому, что при подобной длине филаменты актина и миозина больше не соприкасаются. Таким образом, они в лучшем случае развивают незначительное напряжение.
| 100 - |
|
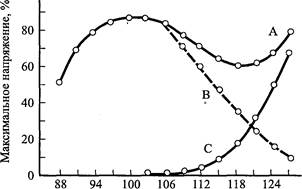
Общее напряжение активной мышцы во время растягивания. Максимальное общее активное напряжение наблюдается в том случае, когда длина мышцы превышает исходную длину или длину в покое в 1,2-1,3 раза. При большей длине общее активное напряжение снижается до тех пор, пока длина мышцы не превышает ее длину в покое примерно в 1,5 раза; в этом случае производство активного напряжения равно нулю. При длине выше 1,3 LQ количество поперечных соединений снижается, степень перекрытия также сокращается, что приводит к уменьшению напряжения. Более того, хотя ПЭК возрастает в своем производстве пассивного напряжения, этот показатель не соответствует снижению активного напряжения сократительных компонентов. Следовательно, величина общего напряжения снижается. При экстремальной длине мышцы пассивное напряжение, генерируемое СЭК, существенно увеличивается, компенсируя тем самым снижение активного напряжения, что приводит к увеличению общего напряжения. Диаграмма длины-напряжения для пассивного растягивания нестимулированной мышцы показана на нижней кривой С. Кривая А, иллюстрирующая общее изометрическое напряжение при стимулировании мышцы различной длины от максимального растяжения до умеренного сокращения, представляет собой сумму активного сокращения (В) плюс напряжение, обусловленное пассивным растягиванием (С). Активное напряжение (кривая В), обусловленное исключительно мышечным сокращением, можно получить, вычитая величину пассивного напряжения С из величины общего напряжения А. Нормальная длина мышцы в покое 100% (рис. 5.13).
Растягивание мыш
цы во время сокраще
ния при большой длине.
Длина в покое, % Ткани при растягивании
Рис. 5.13. Диаграмма длины-напряжения общего и развивают напряжение,
|
|
|
|
|
Дата добавления: 2015-07-02; Просмотров: 466; Нарушение авторских прав?; Мы поможем в написании вашей работы!