КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Чувствительные рецепторы, связанные с растягиванием
|
|
|
|
Три вида рецепторов непосредственно связаны с растягиванием и поддержанием оптимального диапазона движения. Это нервно-мышечные веретена, нервно-сухожильные веретена и суставные механорецепто-ры. Далее мы рассмотрим их структуру, функции и взаимосвязь с растягиванием.
Нервно-мышечные веретена. Основными тензорецепторами мышцы являются нервно-мышечные веретена. Это наиболее подробно изученные проприорецепторы — рецепторы, расположенные в мышцах, сухожилиях и преддверии уха. Их рефлексы связаны с локомоцией или положением тела. В разном количестве их можно обнаружить в большинстве скелетных мышц тела. Особенно много нервно-мышечных веретен содержится в небольших мышцах кисти и глаза. Поскольку мышечные
 Наука о гибкости
Наука о гибкости
волокна заключены в веретенообразную капсулу (оболочку соединительной ткани, окутывающую рецептор), их называют интрафузальными волокнами. Экстрафузальные волокна являются обычными сократительными единицами мышцы. Веретена прикрепляются к обоим концам экстра-фузальных волокон и, таким образом, располагаются параллельно им. Следовательно, при растягивании мышцы веретено также подвергается растягиванию.
Веретено иптрафузалъныхмышечных волокон
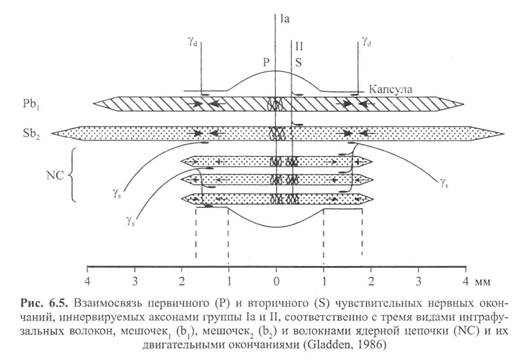
Существует два основных типа интрафузальных волокон: волокна ядерного мешочка и волокно ядерной цепочки. Первые содержат в большом количестве саркоплазму и клеточные ядра в расширенной, напоминающей мешок структуре. Эта несократительная структура находится в центральном, или экваториальном, участке интрафузального волокна. Отсюда и название — интрафузальное волокно ядерного мешочка. На дистальных, или полярных, концах волокна ядерного мешочка находятся исчерченные сократительные филаменты, которые возле веретен прикрепляются к эк-страфузальным волокнам. Не так давно было установлено, что существует два подвида волокон ядерного мешочка. Их назвали мешочек1 и мешочек, (Gladden, 1986; рис. 6.5).
|
Второй тип интрафузального мышечного волокна — волокно ядерной цепочки — более тонкое и короткое, чем волокно ядерного мешочка. Оно содержит всего один ряд ядер, расположенных в виде цепочки по несократительному экваториальному участку. Подобно волокнам ядерного мешоч-
Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиология
ка, полярные концы волокна ядерной цепочки также состоят из исчерченных сократительных филаментов. Их концы нередко соединяются с волокнами ядерного мешочка, которые, в свою очередь, прикрепляются к эндо-мизию экстрафузальных волокон.
Чувствительные нервы веретен
Существует два вида чувствительных нервных (афферентных) окончаний в каждом веретене: первичные и вторичные. Первичные окончания заканчиваются, спиралевидно окружая центральный участок волокна ядерного мешочка и выпуская боковое ответвление к волокну ядерной цепочки. Афферентные аксоны первичных нервных окончаний относятся к волокнам большой группы I. Чтобы отличить эти чувствительные нервные окончания от других, относящихся к этой же группе, большие афференты веретена называют афферентами группы 1а.
Первичные нервные окончания имеют очень низкий порог растяжения и поэтому легко возбуждаются. Их реакции могут быть как фазными (динамическими), так и тоническими. Фазная реакция определяет интенсивность или скорость растягивания путем изменения частоты нейронных импульсов во время растягивания. Частота разрядки быстро увеличивается при начальном растягивании. Затем, по достижении новой длины, она снижается до постоянного уровня, соответствующего новой тонической длине. Следовательно, тоническая реакция определяет длину мышцы. Иными словами, первичные нервные окончания определяют длину и скорость растягивания.
Вторичные нервные окончания образуют разветвленные, напоминающие цветок окончания. Они содержатся в основном в околоэкваториальном сегменте (вблизи экватора) волокон ядерной цепочки. Аксоны вторичных нервных окончаний относятся к афферентным волокнам группы П. В отличие от первичных нервных окончаний, вторичные нервные окончания определяют только тоническую длину мышцы.
Это мотонейроны вызывают соккращение мышечных филаментов в полярных концах интрафузальных мышечных волокон. При сокращении этих концов в центральном экваториальном участке наблюдается пассивное растягивание (там находятся чувствительные нейронные рецепторы). Таким образом, активация гамма-мотонейронов ЦНС может увеличивать количество растяжения, испытываемого чувствительными нервными окончаниями (Banker, 1980).
Двигательные нервы веретена
Двигательные нервы, иннервирующие каждое интрафузальное мышечное волокно в его полярных участках, называются у-эфферентны-ми волокнами, образующими фузимоторную систему. Различают два вида у-аксонов в зависимости от их воздействия на первичные и вторичные чувствительные нервные окончания. Стимулирование так называемого статического у-аксона (ys) может усиливать чувствительность первично-
Наука о гибкости
го нервного окончания к длине, не влияя (или незначительно влияя) на восприятие (чувствительность) скорости. С другой стороны, динамические у-аксоны (yd) могут существенно усиливать восприятие скорости первичного нервного окончания, практически не влияя на чувствительность к длине.
Функция у-системы заключается в контроле чувствительности веретена к растягиванию. Этот процесс сенситизации веретена при помощи у-эф-ферентов известен как у-смещение (Norback и Demarest, 1981). Активация у-мотонейронов приводит к сокращению или укорачиванию интрафузаль-ных мышечных волокон в их полярных участках. При сокращении интра-фузальных волокон экваториальный участок растягивается. Эта центральная деформация приводит к перетягиванию колец в аннуло спиральную часть первичных окончаний и увеличению интенсивности разрядки аффе-рентов групп 1а и П.
Вторая функция у-мотонейронов — поддержание чувствительности веретена во время сокращений мышцы. Когда мышцы укорачиваются, веретено также пассивно укорачивается. Это пассивное сближение двух концов веретена снимает напряжение как с его первичных окончаний (разгрузка веретена), так и со вторичных; такая разгрузка лишает головной мозг информации из веретена относительно изменений мышечной длины. Чтобы предотвратить разгрузку веретена и отрегулировать его чувствительность, у-мотонейроны активируют.
Процесс возбуждения нервно-мышечного веретена
Процесс возбуждения нервно-мышечного веретена можно охарактеризовать следующим образом. Во-первых, к нервно-мышечному веретену прикладывают минимальный растягивающий стимул. Во-вторых, в чувствительном нейронном окончании происходит изменение степени проницаемости, что приводит к производству генерирующего тока (передача заряда через мембрану нервного терминала). Последний, в свою очередь, вызывает деполяризацию, которая называется генератор потенциала. При несколько большей степени растягивания его амплитуда в окончаниях нервно-мышечного веретена увеличивается. Когда деполяризация достигает порога, возникает проведенный потенциал действия. При дальнейшем увеличении степени растягивания может наблюдаться серия проводимых нервных импульсов. Этапы возбуждения нервно-мышечного веретена приведены ниже:
| (1) | (2) | (3) | (4) | (5) |
Стимул Локальное Генератор Локальная деполяри- Проведенный
(растягивание) изменение тока (передача зация (генератор потенциал
проницаемости заряда) потенциала) действия
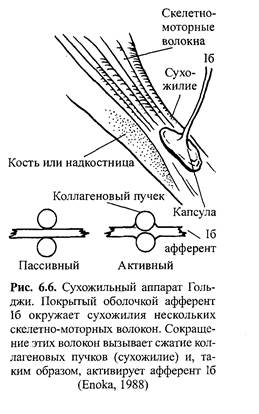
Нервно-сухожильные веретена. Нервно-сухожильные веретена — это чувствительные к сокращению механорецепторы скелетных мышц
Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиология
млекопитающих, иннервируемые быстропроводящими афферентными нервными волокнами группы 16, имеющими большой диаметр (Jami, 1992). Впервые нервно-сухожильные веретена были обнаружены и описаны Гольджи в 1903 году. Ввиду технических трудностей нервно-сухожильные веретена не так хорошо изучены, как нервно-мышечные. Тем не менее их важность с точки зрения развития гибкости нельзя недооценивать.
Местонахождение и структура нервно-сухожильных веретен
Нервно-сухожильные веретена находятся почти исключительно у апоневрозов или мышечно-сухожильных соединений, но не в сухожилиях (рис. 6.6). Так, Пенг, исследовав (цитируется по Bavker, 1974) 1337 рецепторов из различных участков мышц кошек, установил, что 92,4 % их располагались у мышечно-сухожильных соединений и только 7,6 % •— в самом сухожилии.
|
У немлекопитающих нервно-сухожильные веретена представляют собой расположенные вдоль сухожильных пучков не покрытые оболочкой рецепторы (пучки коллагеновых волокон). У млекопитающих же, напротив, они покрыты оболочкой и, как уже упоминалось, располагаются у мышечно-сухожильных или мышечно-апоневротических соединений. Значение этого различия, по мнению исследователей, состоит в следующем. Во-первых, окруженные оболочкой веретена являются более чувствительными к данной величине стимула и более точными в определении и передаче информации в ЦНС. Во-вторых, местонахождение нервно-сухожильных веретен обеспечивает их экстремальную чувствительность к любому изменению напряжения отдельных мышечных волокон, к которым они прикреплены (J.C.Moore, 1984).
| К,и |
Нервно-сухожильные веретена располагаются последовательно, в отличие от нервно-мышечных веретен, расположенных параллельно мышечным волокнам. В исследовании Пенга количество мышечных волокон, соединенных последовательно с нервно-сухожильными веретенами, колеблется от 3 до 50. Каждое нервно-сухожильное веретено, как правило, иннервируется отдельным быстропроводящим афферентным нервным волокном группы 16 (Jami, 1992).
|
|
|
|
|
Дата добавления: 2015-07-02; Просмотров: 760; Нарушение авторских прав?; Мы поможем в написании вашей работы!