КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Температурні адаптації пойкілотермних організмів
|
|
|
|
Температура пойкілотермних змінюється услід за температурою довкілля. Вони переважно эктотермны, вироблення і збереження власного тепла у них недостатньо для протистояння тепловому режиму місць проживання. У зв'язку з цим реалізується дві основні дороги адаптації: спеціалізація і толерантність.
Спеціалізовані види стенотермны, вони пристосовані до життя в таких ділянках біосфери, де коливання температур відбуваються лише у вузьких межах. Вихід за ці межі для них згубний. Наприклад, деякі одноклітинні водорості, що розвиваються в гірських льодовиках на поверхні танучого льоду, гинуть при температурах, що перевищують +(3–5) °С. Рослини дощових тропічних лісів не здатні переносити зниження температури +(5–8) °С. Коралові поліпи живуть лише в діапазоні температур води від +20,5 до +30 °C, т. е. у тропічному поясі океану. Голотурія Elpidia glacialis мешкає при температурі води від 0 до +1 °C і не витримує відхилення від цього режиму ні на один градус.
Інша дорога адаптації пойкілотермних видів – розвиток стійкості кліток і тканин до широкого вагання температур, характерного для більшої частини біосфери. Ця дорога пов'язана з періодичним гальмуванням обміну речовин і переходу організмів в латентний стан, коли температура середовища сильно відхиляється від оптимуму.
Ефективні температури розвитку пойкілотермних організмів. Залежність темпів зростання і розвитку від зовнішніх температур дає можливість розрахувати проходження життєвого циклу видів в конкретних умовах. Після холодового пригноблення нормальний обмін речовин відновлюється для кожного виду при певній температурі, яка називається температурним порогом розвитку, або біологічним нулем розвитку. Чим більше температура середовища перевищує порогову, тим інтенсивніше протікає розвиток і, отже, тим швидше завершується проходження окремих стадій і всього життєвого циклу організму (мал. 13).
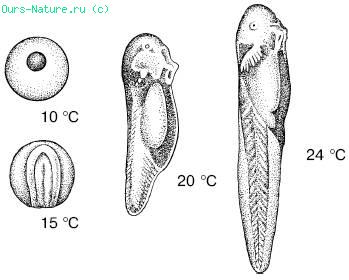
Мал. 13. Стан пуголовків, що розвиваються при різних температурах, через 3 дні після запліднення яйця (по С. А. Зернову, 1949)
Для здійснення генетичної програми розвитку пой‑килотермным організмам необхідно отримати ззовні певну кількість тепла. Це тепло вимірюється сумою ефективних температур. Під ефективною температурою розуміють різницю між температурою середовища і температурним порогом розвитку організмів. Для кожного виду вона має верхні межі, оскільки дуже високі температури вже не стимулюють, а гальмують розвиток.
І поріг розвитку, і сума ефективних температур для кожного виду свої. Вони залежать від історичної пристосованості до умов життя. Для насіння рослин помірного клімату, наприклад гороху, конюшини, поріг розвитку низький: їх проростання починається при температурі грунту від 0 до +1 °C; південніші культури – кукурудза і просо – починають проростати лише при +(8‑10) °С, а насінню фінікової пальми для початку розвитку потрібне прогрівання грунту до +30 °C.
Суму ефективних температур розраховують по формулі
X = (T – C) · t
де X – сума ефективних температур; T – температура довкілля, З – температура порогу розвитку і t – число годинника або днів з температурою, що перевищує поріг розвитку.
Знаючи середній хід температур в каком‑либо районі, можна розрахувати появу певної фази або число можливих генерацій вигляду, що цікавить нас. Так, в кліматичних умовах Північної України може выплодиться лише одна генерація метелика яблуневої плодожерки, а на півдні України – до трьох, що необхідно враховувати при розробці заходів захисту садів від шкідників. Терміни цвітіння рослин залежать від того, за який період вони набирають суму необхідних температур. Для зацвітання мать‑и‑мачехи під Петербургом, наприклад, сума ефективних температур дорівнює 77, кислиці – 453, суниці – 500, а жовтій акації – 700 °C.
Сума ефективних температур, яку потрібно набрати для завершення життєвого циклу, часто обмежує географічне поширення видів. Наприклад, північний кордон лісової рослинності приблизно збігається з липневими ізотермами +(10–12) °С. На північ від тепла для розвитку дерев вже не хапає, і зона лісів змінялася безлісою тундрою.
Розрахунки ефективних температур необхідні в практиці сільського і лісового господарства, при боротьбі з шкідниками, інтродукції нового вигляду і т. п. Вони дають першу, наближену основу для складання прогнозів. Проте на поширення і розвиток організмів впливає безліч інших чинників, тому насправді температурні залежності виявляються складнішими.
Температурна компенсація. Ряд пойкілотермних видів, що мешкають в умовах змінних температур, розвиває можливість підтримувати більш менш постійний рівень обміну речовин в досить широких межах зміни температури тіла. Це явище називається температурною компенсацією і відбувається в основному за рахунок біохімічних адаптацій. Наприклад, у молюсків на побережжі Баренцева морить, таких, як брюхоногие литторины (Littorina littorea) і двостулкові мідії (Mytilus edulis), інтенсивність обміну, що оцінюється по вжитку кисню, майже не залежить від температури в тих межах, з якими молюски зустрічаються щодня під час приливів і відливів. У весенне‑летний період цей діапазон досягає більше 20 °C (від +6 до +30 °C), і в холодній воді їх метаболізм настільки ж інтенсивний, як в теплому повітрі. Це забезпечується дією ферментів, які при пониженні температури міняють свою конфігурацію таким чином, що зростає їх спорідненість до субстрата і реакції протікають активніше.
Інші способи температурної компенсації пов'язані із заміною ферментів, що діють, схожими по функції, але що працюють при іншій температурі (ізоферментами). Такі адаптації вимагають часу, оскільки відбувається инактивация одних генів і включення інших з подальшими процесами збірки білків. Подібна аклімація (зрушення температурного оптимуму) лежить в основі сезонних перебудов, а також виявляється у представників широко поширених видів в різних по клімату частинах ареалу. Наприклад, в одного з видів бичків з Атлантичного океану в низьких широтах Q10 має невисоке значення, а в холодних північних водах зростає при низьких температурах і знижується при середніх. Результатом цих компенсацій є те, що тварини можуть підтримувати відносну постійність активності, оскільки навіть незначне підвищення температури в критичних крапок підсилює змінні процеси. Температурні компенсації для кожного виду можливі лише в певному діапазоні температур, але не вище і не нижче за цю область.
Біохімічні адаптації при всій їх ефективності не представляють головний механізм протистояння несприятливим умовам. Насправді вони є часто «крайнім засобом» і еволюційно виробляються у видів лише тоді, коли неможливі інші способи, фізіологічні, морфо‑анатомические або поведінкові, уникати екстремальних дій без перебудови основного хімізму кліток. Ряд пойкілотермних організмів володіє можливостями часткової регуляції теплообміну, т. е. деякими способами збільшити потрапляння тепла в організм або відвести його надлишок. В основному ці адаптації виникають в багатоклітинних рослин або тварин і в кожній групі мають свою специфіку.
Можливості регуляції температури у пойкілотермних тварин. Найважливіша особливість тварин – їх рухливість, здатність переміщатися в просторі створює принципово нові адаптивні можливості, у тому числі і в терморегуляції. Тварини активно вибирають місця проживання із сприятливішими умовами.
На відміну від рослин, тварини, що володіють мускулатурою, виробляють значно більше власного, внутрішнього тепла. При скороченні м'язів звільняється значно більше тепловій енергії, чим при функціонуванні будь-яких інших органів і тканин, оскільки ККД використання хімічної енергії для здійснення м'язової роботи відносно низький. Чим потужніше і активніше мускулатура, тим більше тепла може генерувати тварину. В порівнянні з рослинами тварини володіють всілякішими можливостями регулювати, постійно або тимчасово, температуру власного тіла.
Пойкілотермні тварини залишаються, проте, як і рослини, эктотермными, оскільки загальний рівень їх метаболізму не настільки високий, аби внутрішнього тепла стало вистачає для обігріву тіла. Наприклад, при температурі +37 °C пустинна ігуана споживає кисню в 7 разів менше, ніж гризуни такої ж величини. Проте деякі з пойкілотермних тварин в стані активності здатні підтримувати температуру тіла вищу, ніж в довкіллі. Наприклад, бабочки‑бражники, ведучі нічний спосіб життя, літають і годуються на квітках навіть при +10 °C. Під час польоту температура грудного відділу підтримується на рівні 40–41 °C. Інші комахи можуть літати в холодному повітрі, заздалегідь розігріваючи свої літальні м'язи для зльоту, наприклад: сарана, джмелі, оси, бджоли, крупні нічні совки і ін. Джмелі збирають нектар навіть при +5 °C, маючи температуру тіла 36–38 °C. При припиненні активності комахи швидко остигають. Генерувати тепло для обігріву можуть в деяких випадках і рептилії. Самка пітона, що обвиває своїм тілом кладку, скорочуючи мускулатуру, здатна підвищувати температуру на 5–6 °C у діапазоні зовнішніх температур від +25 до +33 °C. При цьому вжиток нею кисню зростає майже в 10 разів до граничного для рептилій рівня. У прохолоднішому повітрі змія стає млявою і неактивною.
Основні способи регуляції температури тіла у пойкілотермних тварин – поведінкові: зміна пози, активний пошук сприятливого житла, цілий ряд спеціалізованих форм поведінки, направлених на створення мікроклімату (риття нір, спорудження гнізд і ін.).
Зміною пози тварина може підсилити або ослабити нагрівання за рахунок сонячної радіації. Наприклад, пустинна сарана в прохолодні уранішні години підставляє сонячним променям широку бічну поверхню тіла, а опівдні – вузьку спинну. Ящірки навіть високо в горах в період нормальної активності можуть підтримувати температуру тіла, використовуючи нагрівання прямими сонячними променями і тепло нагрітих скель. По дослідженнях на Кавказі, на висоті 4100 м температура тіла Lacerta agilis часом на 29 °C перевищувала температуру повітря, тримаючись на рівні 32–36 °C. У сильну жару тварини ховаються в тінь, ховаються в норах, щілинах і т. п. У пустелях вдень, наприклад, деякі види ящірок і змій підіймаються на кущі або зариваються в менш нагріті шари піску, уникаючи зіткнення з розжареною поверхнею грунту. Ящірки при необхідності стрімкий перебігають гарячі поверхні лише на задніх ногах, зменшуючи тим самим контакт з грунтом (мал. 16). До зими багато тварин шукають притулки, де хід температур більш згладжений в порівнянні з відкритим житлом. Ще складніші форми поведінки суспільних комах: бджіл, мурашок, термітів, які будують гнізда з добре регульованою усередині них температурою, майже постійною в період їх активності.
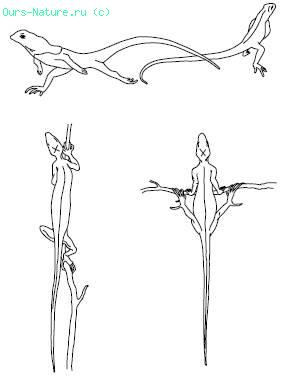
Мал. 16. Поведінка ящірок, що рятуються від розжареної поверхні піску в пустелі
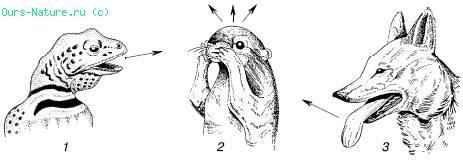
Мал. 17. Випарна терморегуляція у тварин:
1 – ящірка – випар із слизових оболонок при відкритому роті;
2 – антилоповый ховрах – натирання слиною;
3 – койот – випар із слизових оболонок при прискореному диханні
В ряду пойкілотермних тварин ефективно діє і механізм випарної терморегуляції. Жаба за годину при +20 °C втрачає на суші 7770 Дж, що в 300 разів більше за її власною теплопродукции. Багато рептилій при наближенні температури до верхньої критичної починають важко дихати або тримати рот відкритими, підсилюючи віддачу води із слизистих оболонок (мал. 17). Бджоли, що літають в жарку погоду, уникають перегріву, виділяючи з рота краплю рідини, випар якої видаляє надлишок тепла.
Проте, не дивлячись на ряд можливостей фізичної і поведінкової терморегуляції, пойкілотермні тварини можуть здійснювати її лише у вузькому діапазоні температур. Із‑за загального низького рівня метаболізму вони не можуть забезпечити постійність теплового балансу і досить активні лише поблизу від верхніх температурних кордонів існування. Опанування місць проживання з постійно низькими температурами для холоднокровних тварин скрутно. Воно можливе лише при розвитку спеціалізованої криофилии і в наземних умовах доступно лише дрібним формам, здатним використовувати щонайменші переваги мікроклімату.
|
|
|
|
|
Дата добавления: 2014-01-03; Просмотров: 7842; Нарушение авторских прав?; Мы поможем в написании вашей работы!