КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Переваривание и всасывание
|
|
|
|
Гидролиз жиров пищи в просвете тонкого кишечника. Для последующего всасывания ТАГ сначала должны подвергнуться ферментативному гидролизу до свободных жирных кислот (СЖК) и моноацилглицеролов (МАГ). Гидролиз, хотя и в очень малой степени, начинается в желудке под действием кислой липазы. Всасывание жирных кислот инициирует высвобождение холецистокинина и других биологически активных веществ, которые вызывают выделение в составе панкреатического сока липазы и колипазы, секрецию желчных кислот, которые служат для образования эмульгированного жираи мицелл. Панкреатическая липаза катализирует гидролиз ТАГ. Высвобождающиеся при этом СЖК и МАГ всасываются в двенадцатиперстной кишке и проксимальном отделе тонкого кишечника (около 98 %). Всасывание СЖК осуществляют микроворсинки клеток слизистой. Всасывание холестерола (ХС) происходит менее эффективно, чем ТАГ.
Первичные желчные кислоты синтезируются в печени:
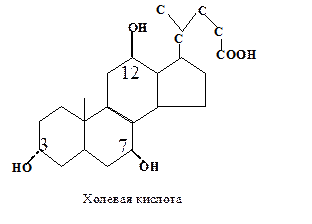
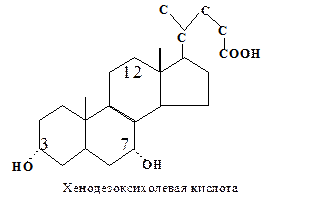
1. Конъюгированные формы первичных желчных кислот. Карбоксильная группа боковой цепи желчных кислот может образовывать амидные связи или с глицином, или с таурином. Это обусловливает их эмульгирующие свойства, так как рК ионной группы боковой цепи ниже, чем у исходной карбоксильной группы.
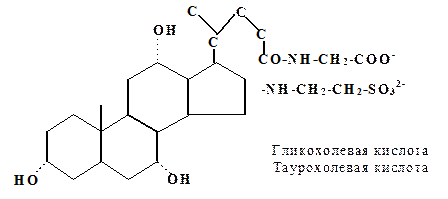
2. Вторичные желчные кислоты (дезоксихолевая, литохолевая) образуются в кишечнике под действием ферментов бактерий, которые катализируют отщепление 7-ОН группы и конъюгированной аминокислоты.
Ресинтез липидов в клетках слизистой тонкого кишечника:

ДЕПОНИРОВАНИЕ И МОБИЛИЗАЦИЯ ЛИПИДОВ ИЗ ЖИРОВЫХ ДЕПО,
ВНУТРИКЛЕТОЧНЫЙ ОБМЕН ЖИРНЫХ КИСЛОТ
Состав липопротеинов. Липопротеины состоят из ядра, в котором находятся триацилглицеролы (ТАГ), эфиры холестерола (ЭХ), и поверхностного монослоя из фосфолипидов (ФЛ), свободного или неэстерифицированного холестерола (СХ) и апопротеинов. Функцией липопротеинов является транспорт липидов. Без этой транспортной формы липиды были бы нерастворимы в плазме крови.
Синтез хиломикронов (ХМ). В клетках слизистой кишечника (энтероцитах) идет эстерификация 2-МАГ и ХС жирными кислотами (ЖК), образуются ТАГ и ЭХ, из которых затем формируются ХМ. Всосавшиеся ЖК активируются, преобразуясь в ацил-КоА. Это происходит в гладком ЭР. Важнейшим структурным компонентом ХМ является белок
(апо В-48). В составе одной частицы ХМ находится одна молекула апо В-48.
ХМ секретируются с базолатеральной поверхности клеток кишечника в лимфу, а оттуда, через грудной лимфатический проток, попадают в систему кровообращения. После того как ХМ попадают в лимфу, они получают от ЛПВП апо С-II, С-III и апо Е.
Катаболизм ХМ. Попадая в систему кровообращения, ХМ быстро подвергаются катаболизму. Уровень ТАГ в плазме крови возрастает через 2 ч после приема пищи, а через
4 ч — постепенно снижается. Время разрушения ХМ зависит от гидролиза ТАГ под действием липопротеинлипазы (ЛПЛ). Кофактором этого фермента является апо С-II. Гидролиз ТАГ приводит к уменьшению размеров ХМ, образуется избыточное количество поверхностных элементов по отношению к объему частиц.
Остатки ХМ разрушаются в печени. Таким образом, в процессе своего катаболизма в кровотоке ХМ поставляют ЖК клеткам периферических тканей (жировой и мышечной), в то время как ХС пищи попадает в печень.
Обмен липопротеинов очень низкой плотности (ЛПОНП), липопротеинов промежуточной плотности (ЛППП) и липопротеинов низкой плотности (ЛПНП). Основной функцией этих липопротеинов является транспорт жирных кислот в составе ТАГ из печени к периферическим тканям, особенно в жировую и мышечную. Для синтеза ЛПОНП в гепатоцитах требуется белок апо В-100 и липиды ЭХ, ТАГ и ФЛ.
Триацилглицеролы для ЛПОНП синтезируются путем эстерификации жирных кислот. Они поступают в гепатоциты из плазмы крови (источником их является, например, липолиз в жировой ткани) или синтезируются de novo в печени. Образование ЛПОНП регулируется после трансляции за счет контроля синтеза апо В-100.
Новосинтезированная частица ЛПОНП содержит одну молекулу белка - апо В-100. Другие белковые компоненты, апо С-II, апо С-III и апо Е, поступают на неё от ЛПВП после того, как ЛПОНП попадают в плазму крови. Они требуются для ускорения метаболизма ЛПОНП.
Обмен ЛПОНП. На эндотелии сосудистой стенки ТАГ в составе ЛПОНП подвергаются действию фермента ЛПЛ. Необходимым кофактором для проявления активности ЛПЛ является апо С-II. ЛПЛ образуется в клетках многих тканей, среди которых наибольшее значение имеют жировая ткань, скелетная и сердечная мышцы, молочная железа во время лактации. ЛПЛ катализирует гидролиз ТАГ в составе ХМ и ЛПОНП до жирных кислот, моноацилглицеролов (МАГ), в результате ЛПОНП превращаются в кровотоке в ЛППП. Фермент проявляет слабую активность по отношению к МАГ и ФЛ.
В жировой ткани синтез ЛПЛ стимулирует инсулин. Тем самым обеспечивается поступление жирных кислот в адипоциты для синтеза и хранения в виде ТАГ. В мышцах ЛПЛ позволяет использовать жирные кислоты для окисления в периоды между приемами пищи, а инсулин подавляет образование этого фермента.
Липопротеины промежуточной плотности (ЛППП). Образование ЛППП происходит из ЛПОНП. Около 75 % ЛППП попадает в печень после связывания апо Е с рецепторами для ЛПНП или рецепторами для апо В/апо Е. В печени они полностью разрушаются. Около 25 % ЛППП в кровотоке подвергается действию другого липолитического фермента, печеночной липазы (ПЛ). Этот фермент катализирует дальнейшее расщепление ТАГ в составе ЛППП. В результате ЛППП превращаются в ЛПНП.
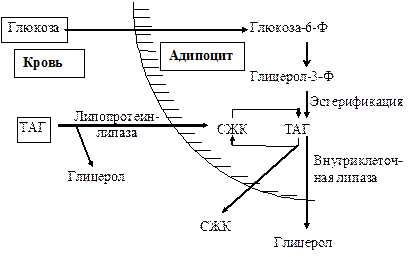
Синтез триацилглицеролов. Клетки большинства тканей, особенно печени и жировой ткани, обладают способностью накапливать ТАГ. Жировая ткань функционально специализируется на хранении (запасании) и мобилизации ТАГ.
Предшественниками для синтеза ТАГ являются глицерол-3-фосфат и активированные жирные кислоты. В печени глицерол-3-фосфат может образовываться или в результате фосфорилирования глицерола, или из глюкозы как промежуточный продукт гликолиза. В жировой ткани единственным источником образования глицерол-3-фосфата является гликолиз.
Вслед за перевариванием пищи в плазме крови увеличивается концентрация глюкозы, инсулина, липопротеинов, богатых ТАГ. Стимулируется активность ЛПЛ для гидролиза ТАГ в составе липопротеинов, и снижается активность жиромобилизующей липазы в жировой ткани. Наряду с этим стимулируется образование ТАГ в жировой ткани. Натощак или при повышенной потребности в энергии во время физической работы, повышении уровня катехоламинов, гормона роста, АКТГ и глюкагона в плазме крови, снижении секреции инсулина эти процессы меняются на противоположные - увеличивается липолиз в жировой ткани и высвобождаются жирные кислоты. Они используются в качестве источника энергии. Глицерол используется для глюконеогенеза.
Схема депонирования и расщепления нейтрального жира в адипоцитах:
Липопротеины низкой плотности (ЛПНП). ЛПНП удаляются из кровотока путем взаимодействия с рецепторами для ЛПНП (другое их название апо В/Е рецепторы). Доля этого процесса в удалении всех ЛПНП составляет 75 %. Остальная часть удаляется с помощью «мусорных» рецепторов, имеющих низкую способность связывания. Этот путь получил образное название «мусорный путь».
Поступление ЛПНП в клетки и их внутриклеточный катаболизм:
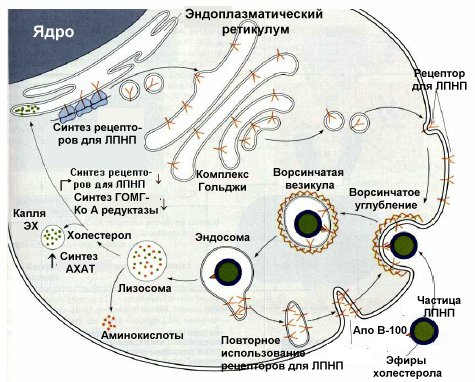
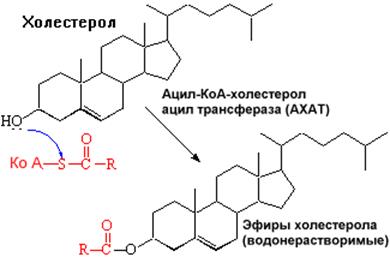
После связывания ЛПНП комплекс рецептор — ЛПНП переносится в клетку посредством эндоцитоза; затем он сливается с лизосомами и разрушается. Внутриклеточное высвобождение холестерола, происходящее таким путем, вызывает следующие эффекты: а) снижает синтез ключевого фермента образования своего, клеточного, холестерола — ГОМГ-КоА редуктазы; б) снижает синтез рецепторов для ЛПНП; в) активирует АХАТ, который катализирует образование из метаболически активной формы СХ метаболически неактивной формы ЭХ.
Схема реакции, катализируемой АХАТ:
В отличие от регуляторного действия рецепторов к ЛПНП на обмен холестерола в клетках, мусорные рецепторы продолжают транспортировать ХС в клетку без торможения по принципу обратной связи. Тем самым макрофаги превращаются в пенистые клетки. Их образование рассматривается как начальный этап атеросклероза.
Метаболизм липопротеинов высокой плотности (ЛПВП). ЛПВП синтезируются в клетках печени и кишечника. Сразу после секреции ЛПВП имеют вид плоских дисков, содержащих ФЛ. Белковым компонентом их является апо А. Из тканей и клеточных мембран на них поступает холестерол. Под действием фермента лецитин-холестерол ацилтрансферазы (ЛХАТ) из СХ и жирной кислоты фосфатидилхолина образуются ЭХ. В результате частицы ЛПВП созревают, принимая форму глобулы. Затем ЭХ переносятся на ЛППП, ЛПНП, обломки ХМ с помощью липидтранспортирующего белка (ЛПБ) или апо D.
|
|
|
|
|
Дата добавления: 2015-07-13; Просмотров: 483; Нарушение авторских прав?; Мы поможем в написании вашей работы!