КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Аэробное окисление углеводов.
|
|
|
|
Распад углеводов в аэробных условиях может идти прямым (aпотомическим или пентозным) путем и непрямым (дихотомическим) путем.
Дихотомическое (греч. dicha - на две части, tome-сечение) окисление углеводов идет по уравнению:
C6H12O6+6O2 ® 6 СО2+б Н2О+686 ккал
Этот путь является основным в образовании энергии. Первые этапы этого пути совпадают с анаэробным окислением глюкозы. Расхождение путей начинается на стадии образования пировиноградной кислоты, которая в животных тканях декарбоксилируется окислительным путем.
Первоначально предполагали, что пировиноградная кислота подвергается окислительному декарбоксилированию с образованием уксусной кислоты по уравнению: СН3—СО—СООН + 1/2 O2 СН3СООН+СО2. Потом было установлено, что уксусная кислота не является промежуточным продуктом при.декарбоксилировании пировиноградной кислоты, и возникло представление, что уксусная кислота существует в «активной» форме. Вскоре было показано, что для утилизации пирувата необходим коэнизм А (КоА). В химическом отношении КоА представляет нуклеотид, в состав которого входит аденозин-31, 51-дифосфат, фосфат, пантотеновая кислота и тиоэтиламин. Коэнзим А участвует в переносе остатка уксусной кислоты — ацетильного радикала (CH3CO-) также и других кислотных (ацильных) радикалов.
Окислительное декарбоксилирование пирувата — процеcc многоступенчатый, осуществляется сложной ферментативной системой, в состав которой помимо пируватдекарбоксилазы, тиаминдифосфата и коэнзима А входят дегидрогеназы с коферментом НАД+, и ФАД, липоевая кислота и ионы магния. В результате окисления пировиноградной кислоты образуются молекула aцетил-КоА («активной» формы уксусной кислоты), два атома водорода (в виде НАДН+Н+) и молекула СО2.
O
÷½
СН3-СО-СООН+НS-КоА+НАД+®СН3--С~S-~КоА+С02+НАДН+ H
Ацетил-КоА
Следующий этап непрямого аэробного окисления глюкозы характеризуется полным окислением ацетил-КоА в цикле Кребса до СО2 и Н2О.
ЦИКЛ ДИ- и ТРИКАРБОНОВЫХ КИСЛОТ (КРЕБСА).
Исследования показали, что дальнейшее окисление ацетил-КоА возможно лишь в присутствии небольших количеств какой-либо ди-карбоновой кислоты. Оказалось, что в начале ацетил-коэнзима А конденсируется с щавелевоуксусной кислотой (СООН—СН2— СО—СООН) с образованием лимонной (трикарбоновой) кислоты. Лимонная кислота является первым продуктом цикла Кребса, поэтому этот цикл иногда называют лимоннокислым.
Образовавшаяся лимонная кислота подвергается далее ряду сложных превращений. И прежде всего, дегидратированию с образованием цис-аконитовой кислоты. Последняя присоединяет молекулу воды и переходит в изолимонную кислоту. Изолимонная кислота подвергается дегидрированию и превращается в щавелево-янтарную, которая декарбоксилируется с образованием µ-кетоглютаровой. µ-Кетоглютаровая кислота подвергается окислительному декарбоксилированию и одновременно дегидрируется, переходя в сукцинил-КоА. Затем сукцинил-КоА превращается в янтарную кислоту. Янтарная кислота дегидрируется, превращаясь в фумаровую. Фумаровая переходит в яблочную, а из яблочной при ее дегидрировании образуется щавелевоуксусная. На этом цикл замыкается. Многие реакции цикла Кребса легко обратимы. Ферменты, катализирующие реакции цикла Кребса
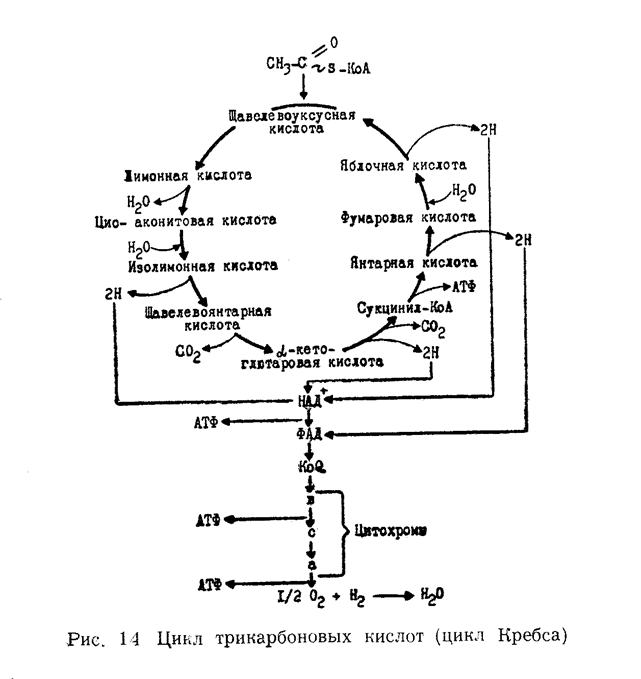 |
сосредоточены в митохондриях. Последовательность реакций цикла Кребса последовательно изображена на суммарной схеме (рис. 7.3).
Из приведенной схемы видно, что в цикле Кребса в результате реакций дегидрирования образуется 4 пары водородных атомов и 2 молекулы СО 2. Освободившийся в ходе процесса окисления водород поступает в цепь биологического окисления и в конечном итоге окисляется молекулярным кислородом с образованием воды и выделением энергии.
При окислении в цикле ди- и трикарбоновых кислот одной молекулы ацетил-КоА образуется 12 молекул АТФ, из которых одиннадцать возникает путем окислительного фосфорилирования, а одна при субстратном фосфорилировании (при превращении сукцинил-КоА в янтарную кислоту).При окислении большинства субстратов в цепи биологического окисления происходит образование 3-х молекул АТФ, тогда как окисление некоторых из них (например, в случае янтарной кислоты) дает 2 молекулы АТФ. Энергетический баланс анаэробного и аэробного окисления глюкозы представляет следующую картину.
1. Две молекулы АТФ — это чистый прирост АТФ при превращениях глюкозы до пировиноградной кислоты в анаэробной фазе.
Глюкоза+2 НАД+ +2 АДФ+2 Фн-® 2 Пируват+2 НАДН+2 АТФ
2. Четыре молекулы АТФ образуются в результате окисления двух молекул НАДН, возникших при дегидрировании двух молекул 3-фосфоглицеринового альдегида (рис. 13). В связи с тем, что эти две молекулы НАДН являются цитоплазматическими, то отдаваемые ими электроны могут включиться в митохондриальную цепь биологического окисления не прямым путем, а с помощью так называемого челночного механизма. Суть этого механизма состоит в том, что сначала цитоплазматический НАДН реагирует с фосфодиоксиацетоном и образует глицерол-3-фосфат.
Фосфодиоксиацетон+НАДН ¬¾ Глицерол-3-фосфат+НАД+
¾®
Г'лицерол-3-фосфат легко проникает через митохондриальную мембрану и окисляется с участием флавинзависимой дегидрогеназы в фосфодиоксиацетон, причем простетическая флавиновая группа восстанавливается.
Глицерол-3-фосфат+Фл. пр. ¾¾® Фосфодиоксиацетон+
4-фл. пр. Н2. Фосфодиоксиацетон выходит из митохондрий, а восстановленный флавопротеид (Фл. пр. Н2) передает приобретенные электроны в цепь биологического окисления, обеспечивая окислительное фосфорилирование только двух молекул АДФ.
3. Шесть молекул АТФ возникают в процессе окислительного де-карбоксилирования двух молекул пировиноградной кислоты, образовавшихся в анаэробной.фазе из одной молекулы глюкозы.
4. При полном окислении двух молекул ацетил-КоА в цикле Кребса возникает 24 молекулы АТФ. В итоге полного аэробного окисления одной молекулы глюкозы синтезируется 36 молекул ЛТФ. При анаэробном гликолизе (брожении) на одну молекулу глюкозы образуется всего 2 молекулы АТФ. Таким образом, «выход» энергии, запасаемой в виде АТФ при кислородном распаде глюкозы, в 18 раз больше, чем при анаэробном.
ПЕНТОЗО-ФОСФАТНЫЙ ПУТЬ ОКИСЛЕНИЯ УГЛЕВОДОВ.
Существует еще один путь аэробного окисления углеводов, который имеет значение лишь в некоторых органах и тканях (жировой ткани, печени, эритроцитах, молочной железе, коре надпочечников, половых железах). Этот путь называется пентозофосфатным (пентозным, прямым или гексозомонофосфатным).
В отличие от непрямого (гексозодифосфатного) пути окисления углеводов в пентозном цикле не проходит стадии образования фруктозе-16-дифосфата, фосфотриоз и т. д., т. е. не происходит фосфорилирования глюкозомонофосфата. Последний в пентозном цикле подвергается прямому окислению с образованием в конечном итоге СО2 и пентозофосфата.
Значение пентозного цикла состоит, во-первых, в том, что является основным поставщиком восстановленного НАДФ + (НАДФН), необходимого для разнообразных синтетических процессов. Во-вторых, этот цикл обеспечивает организм пентозами. В третьих, энергетическая ценность цикла весьма велика — равна 36 мол АТФ.
7.5. ФОТОСИНТЕЗ
Фотосинтез - это синтез органических веществ (прежде всего углеводов) из углекислого газа и воды, происходящий за счет энергии света. Фотосинтез является основным источником образования органических веществ на Земле и единственным источником кислорода.
Процесс фотосинтеза в растениях можно выразить следующим
свет
суммарным уравнением: 6С02+6Н20 ¾® С6 Н 12О6+6 O2
хлорофилл
фотосинтез происходит внутри специализированных органелл- пластидах (хлоропластах), которые содержат в себе хлорофилл.
Хлорофилл — сложное гетероциклическое соединение, содержащее четыре перрольных кольца, образующих порфириновое ядро. Перрольные кольца связаны двумя основными и двумя дополнительными валентностями с атомом магния. Хлорофилл благодаря наличию в его структуре сопряжённых двойных связей способен поглощать световую энергию, переходя в активное состояние. Аккумулирование солнечной энергии хлорофиллом является начальным этапом фотосинтеза, который можно изобразить следующим образом:
Хл + hv —® Хл+
Хлоро- Квант Хлорофилл
филл света возбужденный
На втором этапе фотосинтеза возбужденный хлорофилл, обогащенный энергией за счет поглощения светового фотона, отдаёт энергию на разложение воды (фотолиз воды). В результате образуются активные радикалы Н и ОН. Радикалы ОН дают перекись (ОН)2, распад которой приводит к образованию кислорода.
Хл+ +Н20 —® Хл+Н+ОН
ОН ¾® (ОН)2 ——® H2O+1/2O2
Следующий (третий) этап состоит в переносе водорода воды
через цепь переносчиков на НАДФ с образованием НАДФН. Одновременно происходит за счет переноса электронов, индуцируемого светом, синтез АТФ из АДФ и неорганического фосфата, т. е так называемое фотосинтетическое фосфорилирование.
Разобранные выше этапы являются световой фазой фотосинтеза. В реакциях этой фазы принимает участие поглощенная хлорофиллом световая энергия. Световая фаза приводит к образованию молекулярного кислорода, НАДФН и АТФ.
Следующая фаза фотосинтеза — темновая. Реакции в этой фазе происходят без участия света. В темновой фазе фотосинтеза НАДФН и АТФ используются для восстановления углекислого газа до углевода.
ГЛАВА 8. ОБМЕН ЛИПИДОВ
8.1. РОЛЬ ЛИПИДОВ В ПИТАНИИ
Жиры, так же, как и углеводы, являются важным источником энергии. Кроме того, они выполняют специфические функции благодаря содержащимся в них незаменимым ненасыщенным кислотам: линолевой, линоленовой, арахидоновой, называемых витамином F. Содержатся эти кислоты в основном в растительных маслах. Поэтому растительные масла являются незаменимой составной частью пищи. В жирах содержатся жирорастворимые витамины — А, Д, К, Е, необходимые для нормального развития организма. Богаты витаминами коровье масло, особенно из молока летнего периода, рыбий жир, жир внутренних органов.
Вышесказанное дает основание заключить, что жиры являются неотъемлемой составной частью пищи. Жиры не могут быть заменены другими веществами, хотя и равноценными по калорийности. Суточная потребность человека в жирах составляет в среднем 90 г. При тяжелой физической работе содержание жира в суточном рационе должно быть увеличено. Однако чрезмерное потребление жира ведет к нарушению жирового обмена, к заболеваниям печени.
8.2. ПЕРЕВАРИВАНИЕ ЖИРОВ В ЖЕЛУДОЧНО-КИШЕЧНОМ ТРАКТЕ
Расщепление жира на глицерин и высшие жирные кислоты осуществляется под влиянием фермента липазы. Для воздействия липазы на жир необходимо его предварительное эмульгирование, достигаемое путем перемешивания в кишечнике пищевой кашицы с желчью.
В ротовой полости жиры не подвергаются химическим изменениям. В желудке присутствует липаза, однако ее активность невелика из-за отсутствия условий, необходимых для эмульгирования жира. В желудке гидролизуются только эмульгированные жиры — жиры молока и яичного желтка. В основном переваривание жира происходит в кишечнике и в первую очередь в двенадцатиперстной кишке, куда по протокам попадают вместе с желчью соли желчных кислот, обладающие мощным эмульгирующим действием.
Желчные кислоты образуют тончайшую пленку на жировых каплях, которая препятствует слиянию отдельных капелекжира вболее крупные капли. Это приводит к резкому увеличению поверхности соприкосновения жира с ферментом липазой и, следовательно, скорости гидролитического распада жира. К желчным кислотам относятся холевая, дезоксихолевая и другие. По своему строению они близки к холестерину. В желчи эти кислоты образуют с глицином (гликоколлом) или таурином парные соединения — глико- или таурохолевую, глико- или тауродезоксихолевую и другие желчные кислоты, присутствующие в виде натриевых солей.
В клетках кишечного эпителия из продуктов гидролиза пищевых жиров вновь ресинтезируются жиры, или липоиды, специфичные для данного вида животных. Синтезированные липиды транспортируются в жировые депо. При необходимости из жировых депо жиры могут переходить в кровь и использоваться тканями в качестве энергетического материала.
МЕХАНИЗМ ОКИСЛЕНИЯ НЕЙТРАЛЬНОГО ЖИРА В ТКАНЯХ
Поступивший в клетки нейтральный жир под действием тканевых липаз расщепляется на глицерин и высшие жирные кислоты. В дальнейшем жирные кислоты и глицерин окисляются в тканях на СО2 и H2O, при этом освобождающаяся энергия накапливается в макроэргических связях АТФ.
ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ В ТКАНЯХ. В основе современных представлений о распаде жирных кислот в тканях лежит теория b-окисления, выдвинутая впервые Кноопом в 1904 г. Согласно этой теории, окисление жирных кислот происходит у углеродного атома, находящегося в b-положении по отношению к карбоксильной группе, с последующим разрывом углеродной цепочки жирной кислоты между a- и b-углеродными атомами. В дальнейшем эта теория была уточнена и дополнена.
В настоящее время установлено, что окислению жирных кислот в тканях предшествует их активация при участии коэнзима А и АТФ. Этот процесс катализируется ферментом тиокиназой.
 |
R—СН2—СООН-+НS—КоА+АТФ
О
÷÷
—>R-СН2—С~S—КоА+АМФ+пирофосфат
Ацилкофермента А
Активированная жирная кислота (ацилкофермента А) подвергается дегидрированию, в результате чего возникает двойная связь между a- и b-атомами углерода. Этот процесс протекает с участием ацилдегидрогеназ, которые в качестве простетической группы содержат ФАД. Затем к ненасыщенной кислоте (a, b-не насыщенному производному ацил-КоА) присоединяется молекула воды и образуется b-гидроксикислота (b-гидроксиацил-КоА). Далее снова происходит процесс дегидрирования с образованием b-кетокислоты (b-кетоацил-КоА). Этот процесс катализируется ацилдегидрогеназами, коферментом которых является НАД+.И на последнем этапе b-кетоацил-КоА, взаимодействуя со свободным КоА, расщепляется на ацетил-КоА и ацил-КоА. Последний укорочен по сравнению с первоначальным на два углерода.
Образовавшийся на последнем этапе ацетил-КоА сгорает в лимоннокислом цикле до СО2 и H2O. Оставшийся ацил-КоА подвергается далее аналогичному превращению до полного окисления. Полное окисление насыщенной жирной кислоты приводит к высвобождению значительного количества энергии. Например, при полном окислении пальмитиновой кислоты образуется 131 молекула АТФ.
На основании современных представлений весь ход процесса b-окисления насыщенных жирных кислот можно изобразить следующим образом:
|
|
|
|
|
Дата добавления: 2014-01-04; Просмотров: 1069; Нарушение авторских прав?; Мы поможем в написании вашей работы!