КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Краткий обзор вегетативной иннервации органов1
|
|
|
|
Иннервация глаза. В ответ на определенные зрительные раздражения, идущие от сетчатки, осуществляются конвергенция и аккомодация зрительного аппарата.
Конвергенция глаз — сведение зрительных осей обоих глаз на рассматриваемом предмете — происходит рефлекторно, сочетанным сокращением поперечно-полосатых мышц глазного яблока. Этот рефлекс, необходимый для бинокулярного зрения, связан с аккомодацией глаза. Аккомодация — способность глаза ясно видеть предметы, находящиеся от него на различных расстояниях, зависит от сокращения мышц глаза — m. ciliaris и т. sphincter pupillae. Поскольку деятельность мускулатуры глаза осуществляется совместно с сокращением его поперечно-полосатых мышц, вегетативная иннервация глаза будет рассмотрена вместе с анимальной иннервацией его двигательного аппарата.
Афферентным путем от мышц глазного яблока (проприоцептивная чувствительность) являются, по данным одних авторов, сами анимальные нервы, иннервирующие данные мышцы (III, IV, VI черепные нервы), по данным других — п. ophthalmicus (n. trigemini).
Центры иннервации мышц глазного яблока — ядра III, IV, и VI пар. Эфферентный путь — III, IV и VI черепные нервы. Конвергенция глаза осуществляется, как указывалось, сочетанным сокращением мышц обоих глаз.
Надо иметь в виду, что изолированных движений одного глазного яблока вообще не существует. В любых произвольных и рефлекторных движениях всегда участвуют оба
 Афферентные пути вегетативной иннервации даны в основном по Mitchell (1957).
Афферентные пути вегетативной иннервации даны в основном по Mitchell (1957).
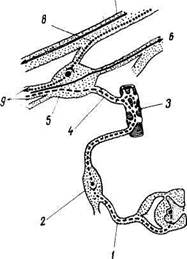
Рис. 343. Ресничный узел (схема).
|
1 — г. commuriicans albus; 2 — gangl. cervicale superius; 3 — a. ophthalmica; 4-х. sympathicus от gangl. ciliare; 5 — gangl. ciliare; 6 — n. nasociliaris; 7 — n. oculomotorius; 8 — radix oculomotoria (парасимпатические преганглионарные волокна); 9 — nn. ciliares breves.
глаза. Эта возможность сочетанного движения глазных яблок (взора) обеспечивается особой системой волокон, связывающей между собой ядра III, IV и VI нервов и носящей название медиального продольного пучка.
Медиальный продольный пучок начинается от ядра в ножках мозга, соединяется с ядрами III, IV, VI нервов при помощи коллатералей и направляется по мозговому стволу вниз в спинной мозг, где заканчивается, по-видимому, в клетках передних рогов верхних шейных сегментов. Благодаря этому движения глаз сочетаются с движениями головы и шеи.
Иннервация гладких мышц глаза — m. sphincter pupillae и т. ciliaris происходит за счет парасимпатической системы, иннервация m. dilatator pupillae — за счет симпатической. Афферентными путями вегетативной системы являются п. oculomotorius и п. ophthalmicus.
Эфферентная парасимпатическая иннервация. Преганглионарные волокна идут из добавочного ядра глазодвигательного нерва (мезэнцефалический отдел парасимпатической нервной системы) в составе п. oculomotorius и по его radix oculomotoria достигают ganglion ciliare (рис. 343), где и оканчиваются. В ресничном узле начинаются постганглионарные волокна, которые через nn. ciliares breves доходят до ресничной мышцы и сфинктера зрачка. Функция: сужение зрачка и аккомодация глаза к дальнему и близкому видению.
Эфферентная симпатическая иннервация. Преганглионарные волокна идут из клеток substantia intermediolateralis боковых рогов последнего шейного и двух верхних грудных сегментов (Суш — ТИц centrum ciliospinale), выходят через две верхние грудные rami commu-nicantes albi, проходят в составе шейного отдела симпатического ствола и оканчиваются в верхнем шейном узле. Постганглионарные волокна идут в составе п. caroticus internus в полость черепа и вступают в plexus caroticus internus и plexus ophtalmicus, после этого часть волокон проникает в ramus communicans, соединяющуюся с п. nasociliaris, и nervi ciliares longi, а часть направляется к ресничному узлу, через который проходит, не прерываясь, в nervi ciliares breves. И те и другие симпатические волокна, проходящие через длинные и короткие ресничные нервы, направляются к дилататору зрачка. Функция: расширение зрачка, а также сужение сосудов глаза.
Иннервация желез — слезной и слюнных. Афферентным путем для слезной железы является
п. lacrimalis (ветвь п. ophthalmicus от п. trigemini), для поднижнечелюстной и подъязыч-
у' ной — п. lingualis (ветвь п. mandibularis от п. trigemini)
 и chorda tympani (ветвь п. intermedius), для околоушной — п. auriculotemporalis и п. glossopharyngeus.
и chorda tympani (ветвь п. intermedius), для околоушной — п. auriculotemporalis и п. glossopharyngeus.
Эфферентная парасимпатическая иннервация слезной
железы. Центр лежит в верхнем отделе продолговатого
мозга и связан с ядром промежуточного нерва (nucleus
salivatorius superior). Преганглионарные волокна идут в
составе п. intermedius, далее п. petrosus major до
ganglion pterygopalatinum (рис. 344). Отсюда начинаются
постганглионарные волокна, которые в составе п. maxilla-
ris и далее его ветви, п. zygoma ticus, через связи
3 en. lacrimalis достигают слезной железы.
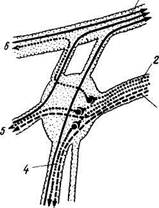
Рис. 344. Крылонебный узел (схема).
1 — п. maxillaris; 2 — п. petrosus major; 3 — п. petrosus profundus; 4 — nn. palatini; 5 — nn. nasales posteriores; 6 — n. zygomaticus.
|
| Hypothalamus |
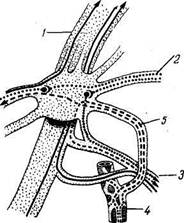
Рис. 345. Ушной узел (схема).
/ — п. mandibularis; 2 — п. petrosus minor; 3 — п. auriculotemporalis; 4 - a. meningea media; 5 — симпатические волокна.
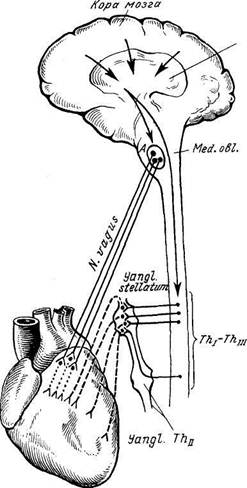
 Рис. 346. Схема иннервации сердца.
Рис. 346. Схема иннервации сердца.
А — ядро сердечных волокон блуждающего нерва в продолговатом мозге; Thj—Th/jj — сегменты спинного мозга, в которых расположены клетки, отдающие симпатические нервные волокна к сердцу. Сплошные линии — ире-ганглионарные волокна блуждающего и симпатических нервов; пунктирные — постгаищо-нарные волокна блуждающего нерва в сердце; прерывистые линии — постганглионарные симпатические волокна, идущие к сердцу. Стрелками показано направление влияния коры мозга, передаваемое через гипоталамус на ядра блуждающего нерва и симпатические центры.
Эфферентная парасимпатическая иннервация подиижнечелюстной и подъязычной жело. Преганглионарные волокна идут от nucleus salivatorius superior в составе п. intermedius, далее chorda tympani и п. lingualis до ganglion submandibulare, откуда начинаются поспан-глионарные волокна, достигающие желез.
Эфферентная парасимпатическая иннервация околоушной железы. Прегапглионарные волокна идут от nucleus salivatorius inferior в составе п. glossopharyngeus, далее п. tym-panicus, п. petrosus minor до ganglion oticum (рис. 345). Отсюда начинаются постганглионарные волокна, идущие к железе в составе п. auriculotemporalis. Функция: усиление секреции слезной и названных слюнных желез; расширение сосудов желез.
Эфферентная симпатическая иннервация всех названных желез. Преганглионарные волокна начинаются в боковых рогах верхних грудных сегментов спинного мозга и заканчиваются в верхнем шейном узле симпатического ствола. Постганглионарные волокна начинаются в названном узле и доходят до слезной железы в составе plexus caroticus internus, до околоушной — в составе plexus caroticus externus и до поднижнечелюстной и подъязычной желез — через plexus caroticus externus и затем через plexus facialis. Функция: задержка отделения слюны (сухость во рту); слезотечение (влияние не резкое).
Иннервация сердца (рис. 346). Афферентные пути от сердца идут в составе п. vagus, а также в среднем и нижнем шейных и грудных сердечных симпатических нервах. При этом по симпатическим нервам проводится чувство боли, а по парасимпатическим — все остальные афферентные импульсы.
Эфферентная парасимпатическая иннервация. 1 Ipei англионарные волокна начинаются в дорсальном вегетативном ядре блуждающего нерва и идут в составе последнего, его
сердечных ветвей (rami cardiaci n. vagi) и сердечных сплетений (см. иннервацию сердца) до внутренних узлов сердца, а также узлов околосердечных полей. Постганглионарные волокна исходят от этих узлов к мышце сердца. Функция: торможение и угнетение деятельности сердца; сужение венечных артерий.
Эфферентная симпатическая иннервация. Преганглионарные волокна начинаются из боковых рогов спинного мозга 4 — 5 верхних грудных сегментов, выходят в составе соответственных rami communicantes albi и проходят через симпатический ствол до пяти верхних грудных и трех шейных узлов. В этих узлах начинаются постганглионарные волокна, которые в составе сердечных нервов, nn. cardiaci cervicales superior, medius et inferior и nn. cardiaci thoracici, достигают сердечной мышцы. Перерыв осуществляется только в ganglion stellatum. Сердечные нервы содержат в своем составе преганглионарные волокна, которые переключаются на постганглионарные в клетках сердечного сплетения. Функция: усиление работы сердца (это установил И. П. Павлов в 1888 г., назвав симпатический нерв усиливающим) и ускорение ритма (это впервые установил И. Ф. Цион в 1866 г.), расширение венечных сосудов.
Иннервация легких и бронхов. Афферентными путями от висцеральной плевры являются легочные ветви грудного отдела симпатического ствола, от париетальной плевры — nn. intercostales и п. phrenicus, от бронхов — п. vagus.
Эфферентная парасимпатическая иннервация. Преганглионарные волокна начинаются в дорсальном вегетативном ядре блуждающего нерва и идут в составе последнего и его легочных ветвей к узлам plexus pulmonalis, а также к узлам, расположенным по ходу трахеи, бронхов и внутри легких. Постганглионарные волокна направляются от этих узлов к мускулатуре и железам бронхиального дерева. Функция: сужение просвета бронхов и бронхиол и выделение слизи.
Эфферентная симпатическая иннервация. Преганглионарные волокна выходят из боковых рогов спинного мозга верхних грудных сегментов (Гйц ~ Thyd и проходят через соответствующие rami communicantes albi и симпатический ствол к звездчатому и верхним грудным узлам. От последних начинаются постганглионарные волокна, которые проходят в составе легочного сплетения к бронхиальной мускулатуре и кровеносным сосудам. Функция: расширение просвета бронхов; сужение.
Иннервация желудочно-кишечного тракта (до сигмовидной кишки), поджелудочной железы, печени. Афферентные пути от указанных органов идут в составе п. vagus, п. splanchnicus major et minor, plexus hepaticus, plexus coeliacus, грудных и поясничных спинномозговых нервов и в составе п. phrenicus.
По симпатическим нервам передается чувство боли от этих органов, по п. vagus — другие афферентные импульсы, а от желудка — чувство тошноты и голода.
Эфферентная парасимпатическая иннервация. Преганглионарные волокна из дорсального вегетативного^ ядра блуждающего нерва проходят в составе последнего до терминальных узлов, находящихся в толще названных органов. В кишечнике это клетки кишечных сплетений (plexus myentericus, submucosus). Постганглионарные волокна идут от этих узлов к гладким мышцам и железам. Функция: усиление перистальтики желудка, расслабление сфинктера привратника, усиление перистальтики кишок и желчного пузыря, расширение сосудов. В составе блуждающего нерва имеются волокна, возбуждающие и тормозящие секрецию.
Эфферентная симпатическая иннервация. Преганглионарные волокна выходят из боковых рогов спинного мозга V— XII грудных сегментов, идут по соответствующим rami communicantes albi в симпатический ствол и далее без перерыва в составе nn. splanchnici majores (VI —IX) до промежуточных узлов, участвующих в образовании чревного, верхнего и нижнего брыжеечных сплетений (ganglia coeliaca и ganglion mesentericum superius et inferius). Отсюда возникают постганглионарные волокна, идущие в составе plexus coeliacus и plexus mesentericus superior к печени, pancreas, к тонкой кишке и к толстой до середины colon transversum; левая половина colon transversum и colon descendens иннервируется из plexus mesentericus inferior. Указанные сплетения снабжают мускулатуру и железы названных органов. Функция: замедление перистальтики желудка, кишок и желчного пузыря, сужение просвета кровеносных сосудов и угнетение секреции желез.
К этому нужно заметить, что задержка движений желудка и кишечника достигается также и тем, что симпатические нервы вызывают активное сокращение сфинктеров: sphincter pylori, сфинктеры кишечника и др.
Иннервация сигмовидной и прямой кишки и мочевого пузыря. Афферентные пути идут в составе plexus mesentericus inferior, plexus hypogastricus superior et inferior и в составе nn. splanchnici pelvini.
Эфферентная парасимпатическая иннервация. Преганглионарные волокна начинаются в боковых рогах спинного мозга И —IV крестцовых сегментов и выходят в составе соответствующих передних корешков спинномозговых нервов. Далее они идут в виде nn. splanchnici pelvini до внутриорганных узлов названных отделов толстой кишки и околоорганных узлов мочевого пузыря. В этих узлах начинаются постганглионарные волокна, которые достигают гладкой мускулатуры названных органов. Функция: возбуждение перистальтики
сигмовидной и прямой кишки, расслабление m. sphincter ani interims, сокращение т. detrusor vesicae и расслабление т. sphincter vesicae.
Эфферентная симпатическая иннервация. Преганглионарные волокна идут от боковых рогов поясничного отдела спинного мозга через соответствующие передние корешки в rami communicantes albi, проходят, не прерываясь, через симпатический ствол и достигают ganglion mesentericum inferius. Здесь начинаются постганглионарные волокна, идущие в составе nn. hypogastrici до гладкой мускулатуры названных органов. Функция: задержка перистальтики сигмовидной и прямой кишки и сокращение внутреннего сфинктера прямой кишки. В мочевом пузыре симпатические нервы вызывают расслабление m. detrusor vesicae и сокращение сфинктера мочевого пузыря.
Иннервация половых органов: симпатическая — см. «Симпатический ствол», парасимпатическая — см. «Периферический отдел парасимпатической системы». Иннервация других внутренних органов приводится после их описания.
Иннервация кровеносных сосудов. Степень иннервации артерий, капилляров и вен неодинакова. Артерии, у которых более развиты мышечные элементы в tunica media, получают более обильную иннервацию, вены — менее обильную; v. cava inferior и v. portae занимают промежуточное положение.
Более крупные сосуды, расположенные внутри полостей тела, получают иннервацию от ветвей симпатического ствола, ближайших сплетений вегетативной нервной системы и прилежащих спинномозговых нервов; периферические же сосуды стенок полостей и сосуды конечностей получают иннервацию от проходящих поблизости нервов. Нервы, подходящие к сосудам, идут сегментарно и образуют периваскулярные сплетения, от которых отходят волокна, проникающие в стенку и распределяющиеся в адвентиции (tunica externa) и между последней и tunica media. Волокна иннервируют мышечные образования стенки, имея различную форму окончаний. В настоящее время доказана наличие рецепторов во всех кровеносных и лимфатических сосудах.
Первый нейрон афферентного пути сосудистой системы лежит в спинномозговых узлах или узлах вегетативных нервов (nn. splanchnici, n. vagus); далее он идет в составе кондуктора интероцептивного анализатора (см. «Интероцептивный анализатор»). Сосудодвига-тельный центр лежит в продолговатом мозге. К регуляции кровообращения имеют отношение globus paliidus, таламус, а также серый бугор. Высшие центры кровообращения, как и всех вегетативных функций, заложены в коре моторной зоны головного мозга (лобная доля), а также впереди и сзади нее. Корковый конец анализатора сосудистых функций располагается, по-видимому, во всех отделах коры. Нисходящие связи головного мозга со стволовыми и спинальными центрами осуществляются, по-видимому, пирамидными и экстрапирамидными трактами.
Замыкание рефлекторной дуги может происходить на всех уровнях центральной нервной системы, а также в узлах вегетативных сплетений (собственная вегетативная рефлекторная дуга).
Эфферентный путь вызывает вазомоторный эффект — расширение или сужение сосудов. Сосудосуживающие волокна проходят в составе симпатических нервов, сосудорасширяющие волокна идут в составе всех парасимпатических нервов краниального отдела вегетативной нервной системы (III, VII, IX, X), в составе передних корешков спинномозговых нервов (признается не всеми) и парасимпатических нервов сакрального отдела (nn. splanchnici pel-vini).
ЕДИНСТВО ВЕГЕТАТИВНОЙ И АНИМАЛЬНОЙ ЧАСТЕЙ НЕРВНОЙ СИСТЕМЫ
Необходимо помнить, что вегетативная нервная система есть часть единой нервной системы. Поэтому в целом организме постоянно наблюдается сочетанная деятельность вегетативной и анимальной частей нервной системы с вовлечением центров, находящихся на различных уровнях нервной системы.
Рассмотрим такую сочетанную деятельность на примере регуляции акта мочеиспускания.
В едином акте мочеиспускания участвуют непроизвольные мышцы (m. detrusor vesicae и т. sphincter vesicae), иннервируемые вегетативными нервами, и произвольная (m. sphincter urethrae), иннервируемая анимальными. При этом вытеснитель мочи сокращается, а оба сфинктера расслабляются, хотя каждый из них иннервируется из разных частей нервной системы:
|
|
| Кишечник |
| Легкие \ ~ Печень (капсула) |
| бронхи |
| Сердце |
| Желудок (подмел, жел) |
| Желудок (паджел. мел) |
| печень Почки Тонкие кишки |
| Мочепол орган |
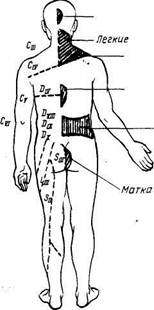
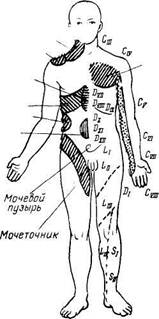
Рис. 347. Зоны отраженных болей (зоны Захарьина — Геда) при заболеваниях внутренних органов.
сфинктер пузыря — из вегетативной, а сфинктер мочеиспускательного канала — из анимальной. Это происходит благодаря наличию общего центра координации в головном мозге, поскольку вегетативная и анимальная части составляют единую нервную систему.
В интеграции анимальной и вегетативной нервных систем большую роль играет лимбическая система.
Зоны Захарьина — Геда. В настоящее время имеются сведения об афферентной иннервации внутренностей анимальной нервной системой, что отмечалось выше. Возможно, этим объясняется давно известный симптом отраженных болей, наблюдаемый в клинике. Заболевания некоторых внутренних органов постоянно сопровождаются отраженными болями в определенных местах кожи. Таковы, например, боли в левой лопатке и левой руке при стенокардии, боли между лопатками при язве желудка, боли в правой подвздошной ямке при аппендиците и др. Эти боли локализуются в определенных кожных сегментах, соответствующих тем сегментам спинного мозга, куда поступают афферентные (чувствительные) волокна из пораженного внутреннего органа. Такие кожные сегменты, или зоны, называются зонами Захарьина — Геда по имени, описавших их авторов. Приводим схему этих зон (рис, 347). Знание зон Захарьина — Геда помогает судить по болям в наружных покровах тела о состоянии органов внутри его полостей. Возможно, что зоны Захарьина — Геда обусловливают воздействие на внутренние органы применяемых в китайской медицине прижиганий или вкалываний игл (акупунктура) в определенные кожные точки.
Сегментарная иннервация органов. Внутриорганные нервы паренхиматозных органов распределяются, как и сосуды, соответственно сегментам органов. Так, в легких различают 10 нервных сегментов. В печени число их весьма варьирует и может достигать восьми. В почке отмечается 5 сегментов иннервации.
ОБЩИЙ ОБЗОР ОСНОВНЫХ ПРОВОДЯЩИХ ПУТЕЙ НЕРВНОЙ СИСТЕМЫ
Как уже отмечалось, интеграция организма в единое целое осуществляется нейрогуморальной регуляцией при ведущей роли нервной системы. Нервная система обеспечивает и единство организма и среды. Рассмотрим морфологическую основу этой интеграции.
В основе деятельности нервной системы лежит рефлекторная дуга. Короткая рефлекторная дуга (см. рис. 264) построена следующим образом. На первом этапе развития центральной нервной системы, когда не было еще головного мозга, рефлекторная дуга замыкалась только в пределах туловищного мозга. Как отражение этого этапа, у человека сохранился собственный аппарат спинного мозга, построенный по принципу трехчленной рефлекторной дуги.
Первый (афферентный, чувствительный) нейрон этой дуги представлен клетками спинномозгового узла, периферические отростки которых идут в составе нервов от органов и тканей, где начинаются рецепторами, а центральные входят в составе задних корешков в спинной мозг. Каждый центральный отросток, войдя в белое вещество спинного мозга, Т-образно разделяется на две ветви — восходящую и нисходящую, от которых в свою очередь отходит несколько боковых веточек (коллатералей). Все эти веточки заканчиваются в задних рогах и substantia intermedia centralis серого вещества нескольких соседних сегментов. Лежащие здесь клетки являются вторым (замыкательным) нейроном простой рефлекторной дуги. Отростки их также делятся на восходящую и нисходящую ветви с боковыми коллатералями, заканчивающимися на клетках передних рогов в пределах нескольких соседних сегментов. Клетки передних рогов составляют третий (эфферентный, двигательный) нейрон; отросток его выходит из спинного мозга в составе передних корешков и далее в составе нервов достигает эффекторов. В результате такого строения простой рефлекторной дуги один чувствительный нейрон вступает в связь с несколькими промежуточными нейронами, а через их разветвления — с еще большим числом двигательных нейронов, вследствие чего раздражение из одной точки тела может передаваться не только на соответственный сегмент, но и на ряд ближайших. Благодаря этому простой рефлекс может стать более распространенным, с вовлечением в ответную реакцию большой группы мышц.
В спинном мозге человека имеются и двучленные дуги, лишенные промежуточного нейрона (см. рис. 351). Примером может служить сухожильный коленный рефлекс, вызываемый постукиванием молоточка по ligamentum patellae при согнутой в колене ноге. В этом случае раздражение передается с сухожильного рецептора на периферический отросток лежащего в спинномозговом ганглии чувствительного нейрона, центральный отросток которого, вступив в составе задних корешков в спинной мозг, достигает серого вещества переднего рога, где и заканчивается на его клетках. Последние образуют второй, двигательный, нейрон, отросток которого в составе переднего корешка и далее мышечного нерва доходит до заложенного в мышцах эффектора. В результате в ответ на постукивание молоточком по сухожилию четырехглавой мышцы бедра наблюдаются ее сокращение и рефлекторное разгибание голени. Двучленная дуга считается молодым приобретением животных, а трехчленный рефлекторный аппарат спинного мозга является филогенетически древним аппаратом. На его базе с возникновением головного мозга стал развиваться более
молодой проводниковый аппарат, связывающий спинной мозг с развивающимся головным.
По мере развития головного мозга возникают и разрастаются двусторонние связи спинного мозга с головным, вследствие чего с возникновением каждого нового этажа последнего увеличивается число связанных с ним афферентных и эфферентных нейронов. Рефлекторная дуга усложняется, так что вместо одного нейрона в каждой ее части появляются цепи нейронов, образующих афферентные и эфферентные проводящие пути. Следовательно, проводящими путями в нервной системе называются тесно расположенные одно возле другого нервные волокна, соединяющие различные отделы ее и объединенные в системы пучков, характеризующиеся общностью строения и функции. С помощью проводящих путей и достигаются единство организма и его связь со средой. Чтобы понять их строение, нужно учитывать основные этапы эволюции центральной нервной системы, последовательное развитие отделов головного мозга (см. «Филогенез»). У человека существуют одновременно и старые, и новые проводящие пути, благодаря которым спинной мозг оказывается связанным со всеми отделами головного мозга (см. рис. 270).
1. С продолговатым мозгом:
а) восходящие — fasciculus gracilis и fasciculus cuneatus, идущие от спинномозговых ганглиев через задние канатики спинного мозга до соименных ядер продолговатого мозга, nucleus gracilis и nucleus cuneatus; б) нисходящие — от ядер, имеющих отношение к равновесию и координации движений, до передних рогов спинного мозга — tractus vestibulospinalis, tractus reticulospinalis и tractus olivospinalis.
2. С мозжечком:
а) восходящие — tractus spinocerebellaris posterior и tractus spinocerebel-laris anterior; они заканчиваются в коре древней части мозжечка, т. е. в черве. Из них задний состоит из отростков клеток nucleus thoracicus задних рогов на своей стороне и входит в мозжечок в составе нижних его ножек — прямой спинно-мозжечковый путь. Передний состоит из отростков клеток substantia intermedia centralis своей и противоположной стороны. Он поднимается до среднего мозга и входит в мозжечок в составе его верхних ножек. Его волокна переходят на противоположную сторону, образуя перекрещенный спинно-мозжечковый путь; б) нисходящие — от мозжечка к спинному мозгу через средний мост и продолговатый мозг.
3. Со средним мозгом:
а) восходящие — tractus spinotectalis, идущий от задних рогов через ствол мозга до крыши (tectum) среднего мозга; на пути он перекрещивается в commissura alba спинного мозга; б) нисходящие (к передним рогам): tractus tectospinalis — от tectum среднего мозга и tractus rubrospinalis — от красного ядра.
4. С передним мозгом:
а) восходящие идут от клеток nucleus proprius задних рогов спинного мозга к таламусу — tractus spinothalamicus lateralis et anterior и перекрещиваются на своем пути в commissura alba спинного мозга; б) нисходящие — tractus thalamospinalis — от таламуса к передним рогам спинного мозга.
С передним мозгом связаны и пути, идущие от нижележащих отделов головного мозга: lemniscus medialis — идет от ядер fasciculus gracilis et cuneatus продолговатого мозга к таламусу, перекрещиваясь на пути в de-cussatio lemniscorum.
5. С развитием коры большого мозга возникают ее связи с ниже
лежащими отделами, над которыми она становится надстройкой. У чело-
века имеются: а) восходящие — tractus thalamocorticalis — от таламуса к коре большого мозга; б) нисходящие — tractus pyramidalis — от коры большого мозга к ядрам черепных нервов, заложенным в мозговом стволе — tractus corticonuclearis, и к передним рогам спинного мозга — tractus corticospinalis (pyramidalis). Чем более развита кора большого мозга, тем более развиты и исходящие из нее пирамидные пути, достигающие наивысшего развития у человека соответственно наивысшему развитию у него новой коры.
Кроме названных основных пучков нервных волокон, возникают и другие пути, связывающие отдельные части спинного и головного мозга между собой. Благодаря им устанавливается единство всей нервной системы.
СХЕМА ПРОВОДЯЩИХ ПУТЕЙ НЕРВНОЙ СИСТЕМЫ
Проводящие пути с точки зрения направления проведения импульса могут быть разделены на две большие группы — афферентные и эфферентные. Афферентные проводящие пути составляют среднее звено — кондуктор того или иного анализатора; поэтому часть их будет рассмотрена вместе с соответствующими анализаторами (см. «Органы чувств»).
|
|
|
|
|
Дата добавления: 2014-01-04; Просмотров: 1563; Нарушение авторских прав?; Мы поможем в написании вашей работы!