КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Посмертные изменения рыбы
|
|
|
|
Рыба, извлеченная из воды, погибает от удушья (асфиксии) вследствие чрезмерного накопления в ее крови и тканях продуктов распада органических веществ, в частности гликогена, что приводит к угнетению эритроцитов и утрате ими способности поглощать кислород. На скорость асфиксии влияет также степень физического и нервного утомления рыбы при вылове.
Различают следующие основные стадии в посмертном изменении рыбы: выделение слизи, посмертное окоченение, автолиз и бактериальное разложение.
Отделение слизи. Поверхность тела только что выловленной рыбы обычно покрыта тонким слоем прозрачной слизи, выделяемой железами кожи. У уснувшей рыбы эти железы еще некоторое время продолжают функционировать и выделять слизь, количество которой на поверхности рыбы при этом заметно увеличивается.
В слизи содержится белковое вещество глюкопротеид муцин, поэтому она является хорошей средой для развития микрофлоры. Из слизи микроорганизмы постепенно проникают в кожу, а затем и в мясо рыбы. Прозрачная у только что уснувшей рыбы слизь под действием микроорганизмов постепенно мутнеет и приобретает неприятный сначала кисловатый, а затем гнилостный запах.
Неприятный запах слизи еще не является признаком порчи, так как слизь легко удаляется при мойке рыбы.
Окоченение, автолиз и бактериальное разложение—в принципе не отличаются от тех же процессов, протекающих в мясе. Специфической особенностью автолиза рыбы является то, что ферментный гидролиз у нее более активно протекает в мышечной ткани, имеющей бурый цвет, что обусловлено более активными протеолитическими и липолитическими ферментными системами, чем в светлой мышечной ткани.
|
|
|
Окоченение. Посмертное окоченение характеризуется затвердением мускулатуры, которое по времени наступает у большинства видов, начиная от головы и заканчивая хвостом рыбы.
Период посмертного окоченения рыб включает отдельные этапы: начало окоченения, начало полного окоченения, максимум окоченения, начало расслабления, конец расслабления. По внешнему проявлению и химизму процессов посмертное окоченение практически идентично двигательному сокращению живой мышцы. Молекулярный механизм сокращения мышц основан на взаимодействии миозиновых и актиновых филаментов.
Изменения, происходящие при окоченении, проявляются разнообразно и отражаются на качестве рыбы. При посмертном окоченении происходит снижение эластичности мяса рыбы (до 20 % первоначальной величины).
Свежая рыба сразу после убоя и до появления начала окоченения характеризуется большим углом прогиба. С нaчaлом нaстyплeния окоченения угол прогиба подобно эластичности резко уменьшается. Расслабление мышечного волокна сопровождается повышением угла прогиба.
Сокращение мышечной ткани при окоченении происходит и в отделенной от скелета мышце, но в отличие от закрепленной мышцы сопровождается изменением ее размеров и формы (рис. 26), что имеет значение при производстве филе из рыбы до наступления в ее тканях процессов посмертного окоченения. Сокращение филе рыбы в период окоченения составляет 20—30 % его длины. Степень сокращения зависит от вида рыбы, части тела, откуда взята мышечная ткань, температуры тела и других факторов.
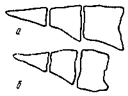
Рис. 26. Размеры и форма различных участков мышц рыбы
до (а) и после (б) наступления посмертного окоченения
Распад гликогена в начальный период окоченения приводит к образованию молочной кислоты и уменьшению рН. Он происходит необязательно до полного расходования гликогена, а в основном до рН 5,4—5,5, при котором инактивируется ферментативная система, обеспечивающая образование молочной кислоты. Однако по количеству оставшегося в мышцах рыб гликогена нельзя судить о глубине прошедших процессов посмертного изменения, так как его количество подвержено исключительно большим колебаниям. Это зависит от обстоятельств, связанных с ловом, способом умерщвления рыбы, ее физиологическим состоянием, определяемым, в свою очередь, факторами внешней среды: температурой, сезонными колебаниями в интенсивности питания, а также периодами воспроизводства и миграции.
|
|
|
По мере развития посмертных изменений рН вновь увеличивается, превосходя первоначальные значения. Несмотря на одинаковый характер изменения рН в посмертный период, имеются особенности, зависящие от исходных значений этого показателя. В большинстве случаев рН мяса неутомленных рыб сразу после их убоя составляет 7,05—7,35, реже 6,2—6,5. В то же время имеются и исключения: рыбы с плоским телом и некоторые другие имеют минимальные величины рН свежего мяса -5,5. Рыбы с минимальными величинами рН сохраняют качество в течение более длительного периода, чем рыбы с более высокими значениями рН.
Гораздо большее значение, чем распад гликогена и образование молочной кислоты, в период посмертного окоченения рыб имеет гидролиз АТФ. Окоченение мышечных волокон начинается, когда примерно 15 % АТФ гидролизовано.
У высокоактивных рыб (тунцов, некоторых окуней) в период после асфиксии тепло генерируется быстрее, чем отводится, и температура тела заметно повышается, в результате чего могут быть ускорены ферментативные процессы в теле рыбы.
Посмертные изменения независимо от вида рыбы проходят в одинаковой последовательности. Однако на время наступления отдельных его периодов и продолжительность их существенно влияют различные факторы: вид рыбы, размер ее, наличие стрессового состояния перед гибелью, способ умерщвления рыбы, температура хранения, способ охлаждения. Продолжительность периодов посмертных изменений в морских рыбах (у рыб, убитых сразу после вылова и хранившихся во льду) составляет: продолжительность от убоя до наступления посмертного окоченения 1—7 ч, продолжительность окоченения 30—120 ч; максимум окоченения приходится на 5—22 ч. У пресноводных рыб продолжительность от гибели до начала окоченения составляет 24—48 ч, продолжительность окоченения — около 72 ч.
|
|
|
При одинаковых внешних условиях продолжительность окоченения зависит от вида рыбы: у подвижных рыб, обладающих интенсивным обменом веществ, посмертное окоченение наступает и заканчивается раньше, чем у малоподвижных, обладающих замедленным обменом веществ. У рыб с плоским телом окоченение наступает медленнее, а протекает быстрее, чем у рыб с торпедообразной формой тела.
Возраст рыбы, а следовательно, и ее размеры сказываются на продолжительности посмертного окоченения: молодые особи подвергаются окоченению быстрее старых.
У рыбы естественной и искусственной популяций наблюдаются различия в скорости протекания посмертных изменений. В рыбе искусственной популяции (на примере красного морского карася) посмертное окоченение начинается раньше, в ее тканях сразу после засыпания содержится гораздо больше молочной кислоты, чем в рыбе естественной популяции. Гидролиз АТФ и накопление продуктов его распада происходят быстрее также у рыбы, выращенной в искусственных условиях.
На продолжительность отдельных этапов окоченения и процесса посмертных изменений в целом наибольшее влияние оказывает снижение температуры хранения улова: снижение ее, например, с 6-12 до 0°С позволяет увеличить продолжительность окоченения в 2 раза.
Понижение температуры с 20 до 0°С способствует уменьшению степени сокращения тканей рыбы на 25-50%. При этом изменяется и степень разрушения ткани, она более значительна при повышенной температуре. Охлаждение рыбы до наступления окоченения способствует сохранению формы, размеров, прочности мышечной ткани и сохранению связи между отдельными волокнами. Влияет на степень сокращения и тип мускулатуры: более чем в 3 раза сильнее сокращаются при окоченении темные мышцы по сравнению со светлыми. Для темной мускулатуры также характерно уменьшение степени сокращения с понижением температуры. Таким образом, для сохранения высоких значений ВУС следует понижать температуру рыбы сразу после ее вылова до наступления посмертного окоченения.
|
|
|
На степень сокращения мышечной ткани рыб, выловленных тралом, влияет воздействие орудий лова, в частности продолжительность траления и степень наполнения трала. При этом обнаруживается зависимость: чем выше способность к сокращению, тем выше качество сырья. Повреждаемость рыбы может проходить в процессе лова, т. е. во время формирования улова; в процессе подъема улова с глубины к поверхности моря; при подъеме из воды и выливке на палубу; при нахождении рыбы на палубе перед длительным хранением.
Механическая повреждаемость гидробионтов в водорыбной пульпе в процессе формирования улова при специализированном промысле, по-видимому, невелика.
Наиболее сильно травмируется рыба во время подъема трала и выливки улова на палубу. При подъеме трала и особенно в момент отрыва его от поверхности воды сильным нагрузкам подвергаются нижние слои, где рыба мнется, повреждаются кожные покровы ее.
Для большинства видов рыб отрицательно сказывается на качестве сырья длительность траления. Степень снижения качества по времени траления для разных видов рыб неодинакова. Отмечена зависимость между размером рыб и снижением ее качества при тралении в течение различного времени: мелкая рыба подвержена большим изменениям, чем крупная. При этом оказывает влияние и степень активности рыбы в трале. Рыба, качество которой снижается быстрее в трале, быстрее портится и при хранении на палубе. Для некоторых видов рыб характерна неоднородность их качества: в одном улове одновременно встречаются особи совершенно свежие и почти неприемлемые в пищу (сабля-рыба, желтохвост).
Степень повреждаемости рыб зависит не только от их вида, но и от присутствия в уловах крупных особей, способных травмировать основной улов, а также скорости подъема трала. При резком подъеме с глубины рыбы получают тяжелые травмы вплоть до выброса внутренностей, Разрыва мышечной ткани и т. д.
Снижение отрицательного влияния процесса добычи рыбы и других объектов может быть получено при правильной организации и квалифицированном выполнении всех операций лова, совершенствовании орудие и методов лова, например путем создания новых тралов, своевременной переборки пассивных орудий лова, ограничения продолжительности траления и величины улова, а также подбора сетных полотен.
Для некоторых рыб установлены критические значения степени сокращения мышц, ниже которой не рекомендуется использовать их с целью замораживания: для скумбрии 12, ставриды 11, хека 7,5 %.
В настоящее время не существует нормативов на предельно допустимый уровень содержания молочной кислоты в мышцах выловленных рыб, введение которого способствовало бы улучшению качества рыбы в период лова. Летальный порог содержания молочной кислоты в мышцах для вылова тралом пикши известен и составляет 100, для лососевых — 125 мг на 100 г.
Автолиз. Окончание процесса окоченения проявляется в расслаблении ткани, восстановлении микроскопической структуры мышечных волокон (при этом могут наблюдаться и нарушенные волокна). На стадии расслабления происходит повышение рН, ВУС, растворимости белков, что положительно сказывается в целом на качестве мяса рыбы.
Процесс окоченения подготавливает мышечную ткань к автолизу - ферментативному распаду. В окоченении кислая среда привела к облегчению выхода протеолитических тканевых ферментов. В период автолиза основные изменения связаны с гидролизом белков и липидов. Переход рыбы из стадии окоченения в автолиз вызывает увеличение активности пептидгидролаз мышечной ткани в щелочной зоне рН. Это увеличение сопровождается уменьшением активности пептидгидролаз внутренностей.
Автолитические изменения затрагивают мышечные белки, белки соединительной ткани, в том числе и жировой. В рыбе протеолиз начинается с тканей, богатых ферментами: в местах скопления крови, желудочного и панкреатического соков. В результате протеолиза повышается проницаемость оболочек внутренних органов и ферменты проникают в брюшную полость, где контактируют с тканями брюшка, ускоряя проникновение их в мышечную ткань.
Протеолиз вызывает глубокие изменения белков и тканей в целом, размягчая их, расслаивая по септам, ослабляя связь с костями и кожей. В то же время изменения, указанные выше, не приводят к порче рыбы.
Первоначальное гидролитическое расщепление белков рыб связано с действием тканевых ферментов. Под действием эндопептидаз белки расщепляются до пептонов и полипептидов. Затем экзопептидазы гидролизуют их до свободных аминокислот (САК).
Одновременно проходит ферментативное дезаминирование аминокислот под действием дезаминазы с образованием и накоплением аммиака. При этом происходит восстановление Т.МАО в ТМА. В период протеолиза рН тканей недостигает оптимума действия протеолитических катепсинов и ферментов внутренностей рыб и процесс идет сравнительно медленно.
Содержание САК в мышцах рыб в процессе протеолиза увеличивается в 3-7 раз по сравнению с посмертным окоченением, при этом значительно изменяется их качественный состав, зависящий от вида рыб.
У рыб, ведущих подвижный образ жизни, протеолиз наступает раньше, чем у малоподвижных. На скорость наступления протеолиза, как и при окоченении, оказывает влияние состояние рыбы перед гибелью.
Основное влияние на скорость протеолиза, как и на процесс окоченения, оказывает температура.
В мясе рыб, где в значительном количестве содержатся азотистые небелковые вещества, определение границ между протеолизом и бактериальным разложением затруднено. Предотвращению нежелательных изменений, вызываемых протеолизом, способствует удаление внутренностей, тщательная зачистка внутренней полости от почечной ткани, обескровливание тканей.
Изменение липидов по отдельно взятым стадиям посмертных изменений изучено недостаточно. Однако изменения этого компонента тканей рыбы в целом в процессе посмертных изменений прослежены подробно. Они сводятся к двум основным направлениям: ферментативному гидролизу и окислению липидов.
Гидролиз липидов рыб, содержащихся как в мышечной, так и в жировой тканях, проходит под воздействием собственных липолитических ферментов.
Среди липидов, содержащихся в мясе рыбы, наиболее устойчивы в период посмертных изменений нейтральные жиры.
Гидролиз глицеридов проходит под действием липаз до жирных кислот и глицерина. Фосфолипиды под воздействием лецитиназ образуют жирные кислоты, холин и фосфорную кислоту.
В посмертный период происходит изменение жирнокислотного состава липидов: высоконенасыщенные жирные кислоты образуют насыщенные и мононенасыщенные с преобладающим образованием олеиновой кислоты.
Бактериальное разложение. Нарушение структуры мышечной ткани при автолизе способствует проникновению в мясо рыбы микробов и развитию в нем бактериальных процессов. Строго разграничить процессы автолиза и бактериального разложения рыбы практически невозможно. Обычно оба эти процесса проходят одновременно, однако интенсивность их может быть различной в зависимости от температуры и санитарных условий хранения рыбы, численности и видового состава обсеменяющих ее микроорганизмов. При повышенной температуре и антисанитарных условиях хранения рыбы бактериальные процессы в ней протекают очень быстро и преобладают над автолитическими. При пониженной температуре, близкой к 0°С, развитие микробов сильно задерживается и автолитический процесс проявляется более отчетливо.
Под воздействием микроорганизмов происходит глубокий распад белковых и жировых веществ рыбы с образованием ряда дурно пахнущих, а также обладающих токсическими свойствами соединений.
Характерные конечные продукты бактериального распада белков — летучие азотистые основания (аммиак и простейшие амины), летучие сернистые соединения (сероводород, меркаптаны), летучие жирные кислоты (масляная и др.), производные группы индола (индол, скатол).
|
|
|
|
Дата добавления: 2013-12-12; Просмотров: 1519; Нарушение авторских прав?; Мы поможем в написании вашей работы!