КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Порядок Сахароміцетальні - Saccharomycetales
|
|
|
|
VI
V
IV
III
II
I
Справжні гриби, в яких вегетативне тіло представлене розгалуженим клітинним міцелієм (зрідка – міцелієм, що брунькується). Вегетативні клітини з генетично однаковими гаплоїдними ядрами (гомокаріонтичні) або дикаріонтичні, причому гаплоїдна стадія за тривалістю, як правило, переважає. Статевий процес – гаметангіогамія, як виняток – соматогамія. Статеве спороношення – аск з аскоспорами. Монадні стадії повністю відсутні.
За видовим багатством аскомікотові є найчисельнішим відділом грибів. До нього належать біля 50 тис. видів (включаючи 16 тис. видів, що утворюють лишайники). Аскомікотові поширені на всіх континентах та практично в усіх основних біотопах.
Біохімічні особливості та живлення. Біосинтез лізину відбувається грибним шляхом (ААА-шлях). Триптофан в аскомікотових синтезується за допомогою ферментів або першої (у більшості представників), або третьої групи (у сахароміцетів). Іони заліза поглинаються за допомогою сидерамінів.
Живлення виключно осмотрофне.
Гормонами, що здатні ініціювати спороношення, є мікоспорини.
Важливими вторинними метаболітами аскомікотових можуть бути антибіотики, мікотоксини, речовини психотропної дії, речовини-інгібітори розвитку вищих рослин, фітогормони.
Цитологічні ознаки. Клітини аскомікотових вкриті оболонками, які утворені переважно хітином та глюканами, а у сахароміцетів – мананом та глюканами. Оболонки аскомікотових двошарові: зовнішній шар тонкий та електронно-щільний; внутрішній – товстий та електронно-прозорий. Септи між клітинами можуть бути простими (часто – з тільцями Вороніна), мікропоровими або доліпоровими з простими пробками.
Ядерний апарат евкаріотичний. Переважаючий у життєвому циклі тип міцелію складається з гіф, утворених одноядерними клітинами. Проте після статевого процесу з клітини злиття, як правило, розвиваються гіфи, що складаються з дикаріонтичних клітин.
Мітохондрії мають пластинчасті кристти. Комплекс Гольджі відсутній, проте у деяких аскомікотових (зокрема у лабульбеніальних грибів) ендоплазматична сітка відшнуровує цистерни, що нагадують типові диктіосоми.
Джгутикові стадії повністю відсутні.
Вегетативне тіло найчастіше представлене розгалуженим септованим міцелієм, рідше (особливо у сахароміцетів) – псевдоміцелієм.
Окремі гіфи міцелію можуть бути видозмінені, найчастіше – в апресорії та гаусторії. Апресорії є органами прикріплення міцелію до субстрату, вони одноклітинні і нагадують присоски. Гаусторії – видозмінені гіфи, що проникають всередину клітини господаря і абсорбують з неї поживні речовини. До видозмінених гіф також належать перфоруючі гіфи – багатоклітинні утвори, що виконують одразу кілька функцій – прикріплення до субстрату, проникнення в нього та поглинання поживних речовин. Перфоруючі гіфи характерні для багатьох грибів – збудників дерматомікозів тварин і людини.
В аскомікотових спостерігається чимало варіантів видозмін не лише гіф, але й міцелію. Серед останніх найпоширенішими є склероції, строми та плодові тіла. Всі ці видозміни пов’язані з утворенням гіфами щільних переплетень, які називають несправжньою тканиною – плектенхімою.
Склероції – це щільні переплетення міцелію, у стані яких гриб переносить несприятливі умови. Зазвичай у склероціях спостерігається диференціація плектенхіми на кору та внутрішню плектенхіму. Гіфи кори товстостінні, забарвлені меланіном у темний колір. Внутрішні гіфи світлі, тонкостінні. У клітинах склероціїв накопичується велика кількість запасних поживних речовин.
Строми також утворені щільними переплетенням гіф, але їх основна функція – це захист (а інколи – піднесення над субстратом) органів нестатевого та статевого спороношень. Тому анатомічна будова стром складніша і включає захисний шар корових гіф, внутрішні гіфи та органи спороношень – пікніди з конідіями або плодові тіла з асками, які занурені у строми.
Плодові тіла – це також видозміни міцелію, утворені плектенхімою. В аскомікотових розрізняють чотири типи плодових тіл: закрите (клейстотецій), напівзакрите (перитецій), відкрите (апотецій) та несправжнє (псевдотецій). Плодові тіла аскомікотових переважно мікроскопічні (виняток становлять пецицальні гриби).
Справжні плодові тіла утворюються з гаплоїдних гіф, з яких частина надалі розвивається в аскогони та антеридії (після статевого процесу з клітин злиття утворюються аскогенні гіфи, що дають початок аскам з аскоспорами), а решта – у захисті структури органів статевого спороношення – плектенхіму плодового тіла. Вона може повністю обростати аски з аскоспорами (тоді утворюються замкнуті плодові тіла - клейстотеції), обростати статеве спороношення лише частково (в цьому випадку утворюються напівзакриті плодові тіла – перитеції, що зазвичай мають вигляд глечика) або розростатися і підносити аски з аскоспорами над субстратом (вкриті плодові тіла - апотеції). Зовнішній шар плектенхіми плодового тіла видозмінюється у його захисну оболонку – перидій. Із внутрішнього шару утворюються додаткові захисні структури (найчастіше – стерильні парафізи).
Несправжні плодові тіла – псевдотеції – на відміну від справжніх, не мають перидію.
Розмноження. У більшості аскомікотових у життєвому циклі спостерігається два типи розмноження – нестатеве та статеве, внаслідок яких розвиваються відповідно два типи спороношень – нестатеве (анаморфа) та статеве (телеоморфа).
Нестатеве розмноження відбувається вегетативним шляхом (фрагментацією справжнього міцелію, брунькуванням, артроспорами, хламідоспорами) або за допомогою різних типів конідій – алевроконідій та бластоконідій.
Конідії можуть утворюватись на поодиноких конідієносцях, в кореміях та спородохіях, на ложах або в пікнідах.
Статеве розмноження відбувається аскоспорами, які утворюються в асках (сумках) внаслідок статевого процесу.
Статевий процес у більшості представників відділу – це гаметангіогамія, при якій зливаються протопласти двох недиференційованих на гамети гаметангіїв – аскогону та антеридію. Рідше плазмогамія відбувається між аскогоном та вегетативною клітиною міцелію, оскільки антеридій не утворюється. У деяких аскомікотовихї статевий процес – це соматогамія, при якій копулюють або звичайні вегетативні гіфи, або аскоспори (інколи навіть не виходячи за межі аску).
Життєвий цикл та розвиток статевих спороношень. У життєвому циклі аскомікотових розрізняють три головні події, які інколи називають кардинальними: статевий процес, внаслідок якого відбувається злиття протопластів клітин статевих партнерів (плазмогамія). Після нього або одразу, або після певного періоду існування у дикаріонтичному стані гаплоїдні ядра зливаються (каріогамія), утворюючи диплоїдне зиготичне ядро. Воно одразу або після певного періоду існування у диплоїдному стані редукційно ділиться (мейоз), після чого починається утворення статевого спороношення – аску з гаплоїдними аскоспорами. Аскоспори або проростають гаплоїдним міцелієм, на якому утворюються статеві органи, або одразу купулюють, розпочинаючи новий життєвий цикл.
В аскомікотових життєві цикли різноманітні, але найпоширенішими та детально дослідженими є три типи: а) гаплодиплофазні без утворення дикаріонтичних поколінь; б) гаплофазні з партеногенетичним утворенням псевдодикаріотичного покоління; в) гаплофазні з чергуванням гаплоїдного та дикаріонтичного поколінь.
Перший тип характерний для сахароміцетів.
Другий тип досліджений на прикладі тафринальних грибів. Тут гаплоїдні клітини, що утворюються внаслідок брунькування аскоспор після одного з мітозів стають двоядерними. Хоча вони нагадують дикаріон, проте містять два генетично однакових ядра, тобто є псевдодикаріонтичними.
Третій тип життєвого циклу серед аскомікотових є найпоширенішим. На гіфах гаплоїдного міцелію, який є переважаючою вегетативною стадією гриба, розвивається багатоядерні антеридії та аскогони з трихогінами (у грибів з порядку евроціальних трихогіна відсутня). Ядра в гаметангіях гаплоїдні. Аскогон (або його трихогіна) контактує з антеридієм, у місті контакту утворюється кон’югаційний канал, по якому ядра з антеридію переходять в аскогон і утворюють дикаріонтичні пари. Із заплідненого аскогону розвивається дикаріонтичні гіфи, що дають початок аскогенним гіфам, верхівки яких згодом перетворюються на аски з аскоспорами. Такий життєвий цикл притаманний більшості аскомікотових, що мають плодові тіла.
Розвиток аску. У більшості аскомікотових аск розвивається за способом гачка: верхівкова дикаріонтична клітина аскогенної гіфи загинається (утворює т. зв. гачок), причому ядра дикаріону розташовуються у зоні загину. Далі вони одночасно мітотично діляться. Пара ядер різної статі залишається в зоні загину гачка, третє ядро переходить у кінчик гачка, а четверте мігрує до базальної частини клітини. Після цього гачок розділяється двома поперечними перегородками на три клітини – верхівкову, одноядерну, середню дикаріонтичну, базальну одноядерну. Далі верхівкова та базальна клітини гачка зливаються, ядро з верхівкової клітини переходить у базальну, відновлюючи дикаріон. Така клітина надалі може утворювати новий гачок. Середня дикаріонтична клітина гачка розвивається в сумку: вона збільшується в розмірах, ядра дикаріона зливаються, утворюючи зиготичне диплоїдне ядро. Воно одразу редукційно ділиться. Далі зазвичай відбувається один мітоз. Навколо кожного з ядер утворюється цитоплазма, яка вкривається оболонкою. Як наслідок, в аску виникає вісім гаплоїдних аскоспор.
У тафриноміцетів розвиток аску простіший.
У сахароміцетів, які не мають у циклі розвитку дикаріонтичної стадії, спосіб утворення аску є найпростішим – диплоїдна клітина, що утворилась після статевого процесу, одразу або після кількох циклів мітозів редукційно ділиться і перетворюється на аск із чотирма аскоспорами.
Розвиток аскоспор. У молодому аску з ядрами майбутніх аскоспор починається формування їх оболонок. Плазмалеми аскоспор можуть утворюватись двома шляхами: з інвагінацій плазмалеми аску та з інвагінацій мембранного навколоядерного мішка.
Типи асків. Залежно від будови аску, аскоспори з нього звільняються або пасивно (внаслідок ослизнення або автолізу оболонки сумки), або активно викидаються з аску.
Аски, з яких аскоспори звільняються пасивно, називають прототунікатними, а ті, з яких аскоспори викидаються примусово, - еутунікатними. Еутунікатні аски, залежно від будови їх оболонки, поділяють на унітунікатні та бібунікатні.
Унітунікатними називають такі аски, які мають лише одну оболонку.
На верхівках унітунікатних асків часто знаходяться додаткові спеціальні структури, які обумовлюють лише апікальний розрив оболонки сумки, і тим самим визначають напрям відстрілу аскоспор. Сумки, які відкриваються на верхівці кришечкою, називають оперкулятними, а ті, які розриваються щілинами – іноперкулятними.
Бітунікатні аски мають дві оболонки – зовнішню (екзоаск) та внутрішню (ендоаск). На верхівці сумки у просторі між цими оболонками також знаходяться мікрофібрилярне кільце.
Система відділу. В межах відділу виділяють шість класів. До провідних фенотипних ознак цих класів належать: а) наявність справжніх плодових тіл; б) тип асків.
Виділяють наступні класи:
1. Сахароміцети – Saccharomycetes. Плодові тіла відсутні. Аски прототунікатні. Покриви аскоспор утворюються з плазмалеми аску. Специфічна особливість – мананово-глюканові клітинні оболонки.
2. Тафриноміцети – Taphrinomycetes. Плодові тіла відсутні. Аски унітунікатні, проте двошарові. Покриви аскоспор утворюються з плазмалеми аску. Специфічна особливість – здатність аскоспор до брунькування.
3. Сордаріоміцети, або аскоміцети – Sordariomycetes (Ascomycetes). Плодові тіла наявні (клейстотеції, перитеції, апотеції). Аски унітунікатні. Покриви аскоспор утворюються з мембран навколоядерного мішка.
4. Леканороміцети – Lecanoromycetes. Плодові тіла наявні (апотеції, рідше перитеції). Аски унітунікатні, леканорового типу. Покриви аскоспор утворюються з мембран навколоядерного мішка. Специфічна особливість – утворення симбіотичних комплексів із водоростями – лишайників.
5. Локулоаскоміцети, або дотідеоміцети – Loculoacomycetes (Dothideomycetes). Плодові тіла наявні (переважно псевдотеції). Аски бітунікатні. Покриви аскоспор утворюються з мембран навколоядерного мішка.
6. Евроціоміцети – Eurotiomycetes. Плодові тіла наявні (переважно клейстотеції). Аски прототунікатні. Покриви аскоспор утворюються з мембран навколоядерного мішка. Специфічна особливість – відсутність у карпогону трихогіни.
Клас Сахароміцети - Saccharomycetes
Статевий процес - соматогамія. Аски прототунікатні, аскоспори звільняються пасивно. Справжні конідіальні спороношення відсутні.
Типовим порядком є Saccharomycetales.
Ознаки порядку повторюють ознаки класу. Сахароміцетальні гриби є переважно сапротрофами, що мешкають у грунті, на поверхні плодів та на інших субстратах, багатих на прості цукри. Деякі cахароміцети у стадії анаморфи здатні викликати хвороби людей (наприклад, рід кандіда - Саndida).
Рід сахароміцес або дріжджі - Saccharomyces (рис. 1). Представники роду Saccharomyces не мають справжнього міцелію, існують у вигляді поодиноких клітин, розмножуються брунькуванням. У практичній діяльності людини найширше використання мають S. сеrеvisiae - цукрові дріжджі. Вони є збудниками бродіння, їх різні штами застосовуються у хлібопекарській та кондитерській промисловості, а також у пивоварінні та виноробстві.
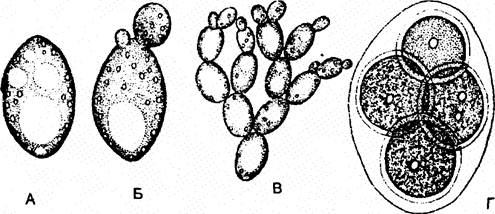
Рис. 1. Saccharomyces cerevisiae: А - вегетативна клітина; Б - клітина, що брунькується; В - псевдоміцелій; Г - аск з аскоспорами
Клас Тафриноміцети - Тарhrinomycetes
Об'єднує гриби, в яких у життєвому циклі наявна коротка гаплоїдна фаза, представлена аскоспорами або міцелієм, що брунькується, та псевдодикаріонтична фаза, протягом якої розвивається розгалужений міцелій, що уражує вищі рослини. Плодові тіла відсутні, аски унітунікатні, проте двошарові, й аскоспори звільняються активно. Клас представляє єдиний порядок Тарhrinales.
|
|
|
|
|
Дата добавления: 2013-12-14; Просмотров: 1476; Нарушение авторских прав?; Мы поможем в написании вашей работы!