КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Олигонуклеотид-направленный мутагенез с использованием плазмидной ДНК
|
|
|
|
Основной недостаток олигонуклеотид-направленного мутагенеза с использованием фага М13 – большое число процедур. Чтобы выделить мутантную форму нужного гена, приходится затратить много времени. В качестве альтернативы было разработано множество других подходов, основанных на применении плазмидных ДНК. Это позволяет обойтись без переноса интересующего исследователя гена из плазмиды в фаговую ДНК, а после завершения мутагенеза – обратно в плазмиду.
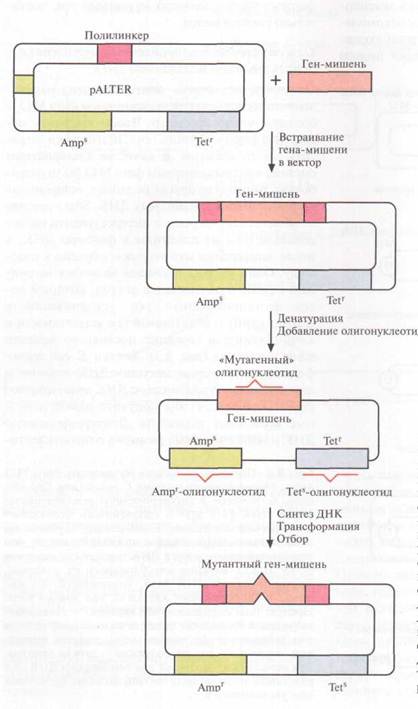
Один из этих подходов включает встраивание ДНК в плазмидный вектор, который несет функциональный ген устойчивости к тетрациклину; и неактивный ген устойчивости к ампициллину; в середине последнего заменен один нуклеотид. Клетки E. Сoli трансформируют вектором, несущим ДНК-мишень, и двуцепочечную плазмидную ДНК денатурируют щелочью с тем, чтобы получить одноцепочечные кольцевые молекулы. Денатурированную ДНК отжигают с тремя разными олигонуклеотидами. Один из них предназначен для внесения изменений в клонированную ДНК-мишень, второй – для устранения мутации в гене устойчивости к ампициллину, третий – для замены одного нуклеотида в гене устойчивости к тетрациклину с тем, чтобы инактивировать этот ген. В реакционную смесь добавляют четыре дезоксирибонуклеозидтрифосфата и ДНК-полимеразу. Гибридизовавшиеся олигонуклеотиды служат затравками для синтеза ДНК, а инактная кольцевая молекула ДНК – матрицей. Одноцепочечные разрывы в новосинтезированной цепи зашиваются с помощью ДНК-лигазы. По окончании синтеза и лигирования (соединения двух молекул ДНК с помощью фосфодиэфирных связей in vitro катализируется ферментом ДНК-лигазой фага Т4) продуктами реакции трансформируют клетки E. сoli. Трансформантов отбирают по признаку устойчивости к ампициллину и чувствительности к тетрациклину. Примерно 90% из них содержат специфическую мутацию в клонированном гене. У остальных трансформантов клонированный ген не был изменен либо потому, что олигонуклеотид не гибридизовался с ним, либо потому, что он вытеснялся в ходе синтеза ДНК. Клетки, несущие мутантный клонированный ген, идентифицируют с помощью гибридизации. Все плазмиды, штаммы, ферменты, олигонуклеотиды (кроме того, который предназначен для изменения клонированного гена), а также буферы продают в наборе, что облегчает работу.
Олигонуклеотид-направленный мутагенез с использованием полимеразной цепной реакции (ПЦР)-амплификации. (умножение числа отдельных генов или группы генов)
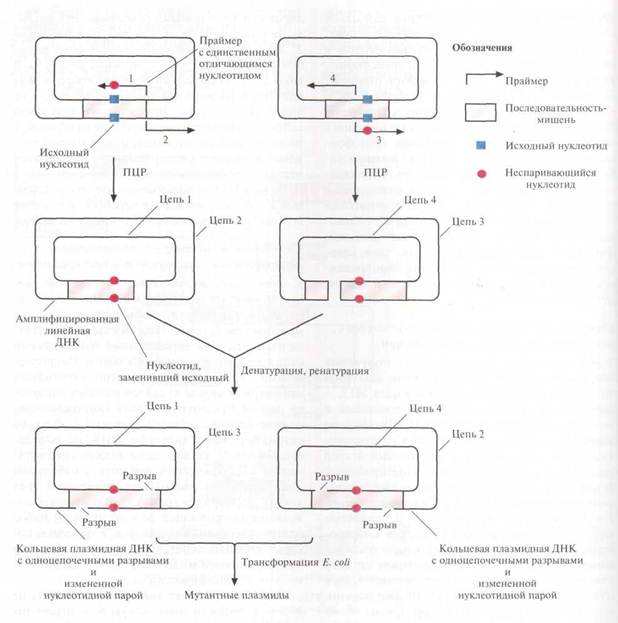
Один из вариантов этого подхода состоит в следующем. Ген-мишень встраивают в плазмидный вектор и помещают препарат в две пробирки. В каждую из них добавляют по два специфических праймера (короткий олигонуклеотид, который гибридизуется с матрицей и служит затравкой при ее копировании) для ПЦР; 1 и 2 в одну пробирку, 3 и 4 – в другую. Праймеры 2 и 4 полностью комплементарны одному из участков клонированного гена или прилегающей к нему последовательности, а 1 и 3 комплементарны другому участку, но содержат один некомплементарный нуклеотид и гибридизуются с разными цепями, так что в результате происходит замена обоих нуклеотидов данной пары. Положение сайтов гибридизации праймеров 1 и 2 в одной пробирке и 3 и 4 – в другой таково, что ПЦР-продукты в разных пробирках имеют разные концы. По окончании ПЦР содержимое пробирок объединяют и проводят денатурацию, а затем ренатурацию (воссоединение цепей двуцепочечной ДНК, разошедшихся при денатурации). Поскольку концы амплифицированных молекул ДНК из двух пробирок неодинаковы, одноцепочечные ДНК из разных пробирок ассоциируют с образованием кольцевых молекул с двумя одноцепочечными разрывами. Эти разрывы репарируются in vivo после трансформации E. сoli. При ренатурации одиночных цепей из одной пробирки образуются линейные молекулы. В клетках E. сoli стабильно поддерживаются в виде плазмид и наследуются только кольцевые, а не линейные молекулы, при этом все они несут сайт-специфическую мутацию.
|
|
|
|
|
Дата добавления: 2014-01-05; Просмотров: 2666; Нарушение авторских прав?; Мы поможем в написании вашей работы!