КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Пути обмена аминокислот
|
|
|
|
В основе различных путей обмена аминокислот в организме лежат три типа общих реакций: реакции по α-аминогруппе, по карбоксильной группе и по радикалу аминокислоты.
Реакции по α-аминогруппе однотипны у всех аминокислот и заключаются в реакциях дезаминирования и переаминирования (трансаминирования).
Реакции по карбоксильной группе тоже однотипны для всех аминокислот и сводятся в основном к декарбоксилированию и образованию аминоациладенилатов.
В отличиеот этого преобразования в радикалахаминокислот исключительно разнообразны и, как правило, уникальны для каждой отдельной аминокислоты.
Часть безазотистого углеродного скелета аминокислот, образовавшихся в результате дезаминирования, подвергается окислительному расщеплению на фрагменты, которые могут включаться затем в лимоннокислый цикл для дальнейшего окисления.
С другой стороны, безазотистые углеродные скелеты аминокислот могут включаться в биосинтетические реакции. Многие аминокислоты (10 аминокислот) после утраты аминогруппы могут превращаться в углеводы. Их называют глюкопластическими (основные – аланин, аспарагиновая и глютаминовая кислоты). Другие три аминокислоты (лейцин, фенилаланин и тирозин) включаются в метаболизм липидов и их именуют кетопластическими аминокислотами.
Пути катаболизма аминокислот сложны и переплетаются со многими другими метаболическими путями, благодаря чему многие промежуточные продукты катаболизма аминокислот служат необходимыми предшественниками для синтеза различных компонентов клетки и биологически активных веществ.
Катаболизм аминокислот у млекопитающих происходит главным образом в печени и несколько слабее в почках.
Остановимся на вышеперечисленных общих реакцияхпоподробнее. В реакциях по α-аминогруппе, связанных с дезаминированием, различают четыре вида дезаминирования аминокислот:
- окислительное дезаминирование;
- восстановительное дезаминирование;
- гидролитическое дезаминирование;
- внутримолекулярное дезаминирование.
Указанные реакции катализируются специфическими ферментами. Наиболее широко распространено окислительное дезаминирование, катализируемое оксидазами аминокислот (их еще называют дегидрогеназами), тогда как остальные виды дезаминирования встречаются редко (лишь у отдельных групп организмов).
Окислительное дезаминирование осуществляется в два этапа. Сначала на первом этапе аминокислота окисляется в иминокислоту при участии специфической оксидазы (дегидрогеназы), где в качестве простетической группы в случае с оксидазой глутаминовой кислоты (глутаматдегидрогеназы) выступает НАД, тогда как для дегидрогеназ (оксидаз) других аминокислот – ФМН (для L-аминокислот) или ФАД (для Д-аминокислот).При этом НАД, ФМНилиФАДвосстанавливаются.
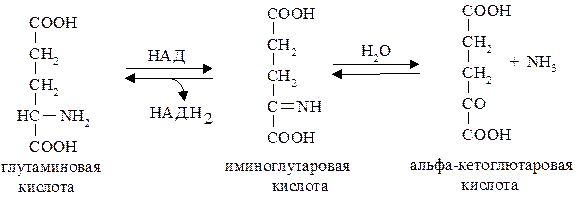 |
В животных тканях Эйлером выявлен высокоактивный при физиологических значениях рН фермент глутаматдегидрогеназа, катализирующая окислительное дезаминирование L‑глутаминовой кислоты. Этот процесс можно представить в виде следующих уравнений.
Надо заметить, чтообе реакции обратимы и такимпутем изα‑кетоглутаровой кислоты и аммиака в организме может образовываться глутаминовая кислота.
Благодаря обратимости этой реакции, обмен глутаминовой кислоты связан с центральным путем катаболизма субстратов – лимоннокислым циклом.
Дегидрогеназы (оксидазы L‑ и Д‑аминокислот), присущие организму в тканях животных и растений, при физиологическом значении рН в общем малоактивны и только дегидрогеназа глутаминовой кислоты проявляет себя высокоактивным ферментом. Поэтому считают, что большинство аминокислот в организме дезаминируются непрямым дезаминированием,так называемым транс-дезаминированием, при котором аминокислоты предварительно переаминируются в основном с α-кетоглутаровой кислотой, а затем образовавшаяся в результате этого глутаминовая кислота подвергается окислительному дезаминированию.
Общий итог переаминирования различных аминокислот состоит в том, что все их аминогруппы собираются в общий фонд в виде одной аминокислоты, обычно в виде глутаминовой кислоты. У некоторых организмов первоначально такой аминокислотой может быть аланин или аспарагиновая кислота, однако в дальнейшем происходит перенос аминогрупп с этих аминокислот на α-кетоглутаровую кислоту с образованием глутаминовой кислоты. Следовательно, независимо от первого этапа переаминирования, конечным акцептором аминогрупп большинства аминокислот служит α-кетоглутаровая кислота, которая превращается в глутаминовую кислоту.
Реакция переаминирования катализируется аминотрансферазами. В качестве кофермента (простетической группы) аминотрансфераз (трансаминаз), катализирующих эту реакцию, служит пиридоксальфосфат, являющийся производным витамина В6.
Реакция переаминирования многостадийна.
На первой стадии ферментативного катализа простетическая группа фермента взаимодействует с аминокислотой, подвергающейся переаминированию. Соединение идет по аминогруппе кислоты и альдегидной группе пиридоксальфермента.
На второй стадии катализа идет таутомерная перегруппировка в образовавшемся комплексе.
В результате последующего гидролиза происходит образование и освобождение кетокислоты и пиридоксальаминофермента.
Далее пиридоксальаминофермент образует фермент-субстратный комплекс с α-кетоглутаровой кислотой.
Полученное соединение подвергается таутомерной перегруппировке, гидролизуется и возникает глутаминовая кислота.
Образовавшаяся глутаминовая кислота подвергается окислительному дезаминированию по вышеописанной схеме с превращением в α-кетоглутаровую кислоту, и весь процесс может возобновиться снова. Полагают, что накопление аминогрупп в форме глутаминовой кислоты в результате процесса переаминирования происходит в цитоплазме клеток, после чего эта глутаминовая кислота с помощью особой системы переноса транспортируется через мембрану внутрь митохондрий. В матриксе митохондрий, с одной стороны, с помощью специфической аспартат-глутамат-трансаминазы происходит перенос аминогруппы с глутаминовой кислоты на щавелево-уксусную кислоту с образованием аспарагиновой кислоты, включающейся в орнитиновый цикл, а, с другой стороны, происходит окислительное дезаминирование глутаминовой кислоты глутаматдегидрогеназой с включением выделившегося аммиака в орнитиновый цикл.
Реакция переаминирования между аминокислотами и α-кетоглутаровой кислотой обратима, поэтому при определенных условиях она служит для синтеза аминокислот из кетокислот и глутаминовой кислоты.
Превращения аминокислот в организме, связанные с реакциямипо их карбоксильной группе, сводятся в основном к декарбоксилированию и образованию аминоациладенилатов.
Декарбоксилирование аминокислот имеет место в тканях животных и растений, а такие у микроорганизмов. Декарбоксилирование, как было сказано выше, служит причиной образования аминов при гниении белков в кишечнике. В тканях декарбоксилирование аминокислот также сопровождается выделением СО2 и образованием аминов (гистамина из гистидина, γ-аминомасляная из глутамина и т.д.).
Поскольку амины обладают высокой физиологической активностью и образуются в качестве продуктов жизнедеятельности их называют биогенными аминами. Реакция декарбоксилирования аминокислот ускоряется специфическими для каждой индивидуальной аминокислоты декарбоксилазами, простетической группой которых служит пиридоксальфосфат. Эти ферменты относятся к классу лиаз.
Все амины проявляют свое физиологическое воздействие при весьма малых концентрациях, поэтому их образование в большой концентрации могло бы представить угрозу для нормальной жизнедеятельности. Однако в животных тканях имеются активные аминоксидазы (моно- и диаминоксидазы), которые окисляют амины в соответствующие альдегиды. Последние окисляются в жирные кислоты и распадаются далее до конечных продуктов путем β-окисления.
Второй характерной реакцией по карбоксильной группеаминокислот является образование аминоациладенилатов при участии специфического фермента (аминоацил-тРНК-синтетазы) и АТФ. Значение этой реакции существенно в механизме биосинтеза белка. Образование аминоациладенилата активирует аминокислоту перед вступлением последней в пептидную связь при биосинтезе белка на рибосомах.
Помимо реакций по аминной и карбоксильной группе известны превращения аминокислот, связанные с реакциями по радикалу, т.е. по той части ее молекулы, которая не принимает участия в формировании хребта полипептидной цепи.
Важнейшими типами превращений аминокислот, протекающими с видоизменением радикалов, являются окислительно-восстановительные реакции, β‑декарбоксилирование, деметилирование и переметилирование, комбинированные реакции, которые ведут к переходу одних аминокислот в другие, а также в другие вещества, в том числе гормоны (например - адреналин).
Таким образом, в процессе превращений аминокислот могут возникнуть соединения, принимающие участие в регуляции обмена веществ. Это подчеркивает ведущую роль белкового обмена в общем обмене веществ в организме.
Итак, в результате превращений аминокислот по аминогруппе, по карбоксильной группе и по их радикалу возникают углекислота (СО2), аммиак (NН3), амины, карбоновые кислоты, кетокислоты и в некоторых случаях достаточно сложные вещества, относящиеся к тем или иным классам органических веществ. Если эти вещества не вовлекаются в процессы синтеза, то все они, за исключением углекислоты и аммиака, в конце концов подвергаются дальнейшей деструкции.
Следовательно, конечными продуктами распада аминокислот являются вода, углекислота и аммиак.
|
|
|
|
|
Дата добавления: 2014-01-05; Просмотров: 4096; Нарушение авторских прав?; Мы поможем в написании вашей работы!