КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Биологические особенности рыб в связи с их особенностями роста
|
|
|
|
С физиологической точки зрения кормление рыб искусственными кормами должно обеспечить необходимое соответствие количества, качества, и свойства пищи с требованиями и потенциальными возможностями роста рыб.
ХИМИЧЕСКИЙ СОСТАВ искусственных кормов РЫБ
Таким образом, биохимический состав природного рациона большинства рыб богат белком, который составляет более половины сухого вещества пищи и является преобладающим источником энергии (около 60%). Углеводы находятся в минимальных количествах.
Как видно, в естественной пище рыб (зоопланктон, зообентос) около 60% всей энергии представлено энергией белка.
На современном уровне развития физиологии кормления животных под сбалансированным рационом понимают такой набор кормовых средств, при котором все потребности организма удовлетворяются без избытков каких-либо питательных веществ.
Сбалансированные рационы должны обеспечивать высокий темп роста рыб при низких затратах корма, высокую устойчивость организма при низкой смертности, способствовать нормальному формированию воспроизводительной системы, получению продукции высокого качества, быть экономичными и т. д.
Поэтому для создания сбалансированных рационов необходимо выполнение широкого круга физиолого-биохимических исследований, направленных, с одной стороны, на изучение потребностей организма рыбы в питательных веществах, а с другой — на изучение питательной ценности кормовых средств, характеризующей их способность удовлетворять потребности организма.
Особое внимание в последний период уделяется аминокислотному питанию. Это связано с тем, что биологическая ценность белковой части корма определяется не столько абсолютным содержанием белка в рационе, сколько соотношением незаменимых и заменимых аминокислот, а также степенью их доступности организму животных.
Коллективом американских исследователей под руководством Хальвера на экспериментальных диетах, содержащих синтетические аминокислоты в пропорциях, аналогичных белку куриного яйца и икры, а такжесвободных эмбрионов чавычи, была последовательно определена степень незаменимости 18 основных протеиногенных аминокислот для чавычи, нерки, кижуча, радужной форели. Показано, что в питании этих рыб незаменимы те же 10 аминокислот, что и у теплокровных животных: аргинин, гистидин, лизин, валин, метионин, триптофан, фенилаланин, лейцин, изолейцин и треонин. Основной симптом недостаточности — задержка роста. Рыбы, не получавшие с пищей а -аланина, аспарагиновой и глютаминовой кислот, цистина, пролина, глицина, серина, тирозина, росли так же хорошо, как и рыбы, получавшие все 18 аминокислот.
Аналогичные результаты были получены позднее для тепловодных рыб (канальный сом, угорь, карп), а также некоторых морских рыб (камбала, морской окунь, морской язык). Показана обязательность наличия впище тех же 10 аминокислот.
Для определения количественной потребности в незаменимых аминокислотах содержание каждой аминокислоты увеличивалось в линейной последовательности. Установлено, что оптимальный рост обеспечивало следующее количество и соотношение незаменимых аминокислот в диете при общем содержании белка в рационе 40% (в процентах к белку): аргинин – 6,0, гистидин – 1,8, лизин – 5,0, валин -3,2 метионин – 4,0, триптофан – 0,5, фенилаланин – 5,1, лейцин – 3,9, изолейцин – 2,2.
Для тепловодных рыб определение количественных потребностей в незаменимых аминокислотах оказалось методически более сложным. Первые эксперименты с молодью карпа, сома и морского леща окончились неудачей. В опытах с угрем удалось определить количественные потребности только в некоторых аминокислотах. Показано, что его потребность в триптофане и метионине была вдвое выше, чем у лососевых.
Так как прямые определения для карпа отсутствовали, М.А. Щербиной была предложена гипотетическая модель потребности в незаменимых аминокислотах. Она была построена на основе расчетного метода Уильямса, - согласно которому наиболее благоприятные для анаболизма (процессы синтеза) соотношения незаменимых аминокислот близки к таковым в тотальном белке организма (или мышечной ткани). Гипотетическая потребность двухлетних карпов в незаменимых аминокислотах при общем содержании белка в рационе 20% составила (в процентах к белку): аргинин – 6,0, гистидин – 2,5, лизин – 7,5, валин - 4,0 метионин – 2,5, фенилаланин – 7,0, лейцин – 7,0, изолейцин – 3,5.
При использовании данных о потребностях в незаменимых аминокислотах следует помнить, что они дают представление лишь о средних величинах и являются ориентировочными. Их можно применять для периода активного роста рыб.
Потребность в белке
Рыбы отличаются высокой потребностью в белке, которая существенно превышает таковую у высших позвоночных. Впервые эта особенность питания была замечена при составлении кормовых рационов в условиях искусственного разведения рыб.
Относительно общего количества белка, необходимого для нормального роста и развития различных видов рыб, до настоящего времени нет строго определенных нормативов. На потребность в белке, существуют различные точки зрения, аргументированные соответствующими экспериментами. Это объясняется не только специфическими особенностями источников белка (животного или растительного происхождения), количественным содержанием и соотношением аминокислот, а также сложностью взаимосвязей между аминокислотами и другими питательными веществами (углеводами, жирами, макро- и микроэлементами, витаминами и т. д.), степенью обеспеченности белковой части кормов энергией и другими факторами.
В ранних работах назывались низкие цифры потребностей лососевых в белке — 14%. Такой уровень поддерживался в пастообразных кормах, т. е. в кормах, содержащих большое количество воды. В пересчете на сухое вещество белок составлял в них около 25% от всей массы корма. В более поздних работах в пастообразные корма лососевым рыбам рекомендовалось вводить до 28% белка, что по сухому веществу составляло уже более 50% (Phillips, Brockway, 1956). В дальнейшем, при появлении сухих гранулированных кормов, многие исследователи подтвердили необходимость высокого уровня белка в рационах форели и лосося разного возраста — 40-60% (Halver et al., 1964; Mann, 1968; Hashimoto, Okaiti, 1968; Phillips, 1970; Cowey, 1975; Канидьев, Гамыгин, 1975a, 1977 и др.), что сначала связывали с хищническим питанием лососевых рыб, но затем выяснилось, что и такая мирная рыба, как карп, также требует для оптимального роста высокого уровня белка в корме — 40-50% для сеголеток и 30-40% — для годовиков карпа (Hashimoto, Okaiti, 1968; Steffens, 1969; Meske, 1973; Sen et al., 1978; Скляров и др., 1981а).
Потребность рыб в высокобелковой пище сложилась исторически и связана с их питанием в природе преимущественно животными организмами. Всякие попытки кормить рыб дешевыми растительными рационами, предназначенными для сельскохозяйственных животных, терпели и терпят неудачу. Применение таких кормов в рыбоводстве не позволяет выявить истинные возможности индустриальных методов разведения рыб, резко снижает рост, истощает иммунную систему,
Согласно экспериментальным данным, полученным на полусинтетических диетах, потребность в белке у форели и различных групп лососевых достаточно близка и составляет в первые четыре недели около 50% сухого вещества диеты. С возрастом и увеличением веса потребность снижается. В период 5—12 недель оптимальное количество белка составляло 40%, а далее потребности могли удовлетворяться меньшим количеством. Сходная зависимость была обнаружена у угря, канального-сома и карпа. Сходные цифры потребности в белке указывались для лосося, чавычи, кумжи, радужной и ручьевой форели, угря, сиговых, карпа, сазана, буффало, амура, толстолобика, канального сомика, леща и других видов рыб. При этом оптимальное содержание белка для молоди было выше, чем для старших возрастных групп рыб.
Доказано, что потребность в протеине меняется в зависимости от:
· физиологического состояния рыб (зрелые особи нуждаются в меньшем количестве протеина, чем молодь),
· от условий окружающей среды (с повышением температуры потребность возрастает, аналогичное влияние оказывает и соленость).
· установлено, что увеличение общей калорийности рационов холодноводных и тепловодных рыб за счет жиров и углеводов оказывает азот-сберегающий эффект, в результате чего значительно сокращается количество протеина — корма, необходимого для получения единицы прироста.
В связи с тем, что большая часть исследований проводилась не с синтетическими диетами, а с кормовыми смесями, составленными из натурального сырья, соотношение и доступность питательных веществ которых существенно различались, количества белка, обеспечившее наилучший рост рыб, колебалось в широких пределах. Так, по данным И. Н. Остроумовой (1974), содержание белка в пределах 30—37% обеспечивает быстрый рост сеголетков форели; по данным А. П. Канидьева и др. для той же возрастной группы содержание белка должно составлять 40—45%, оптимум белка для двухлетков форели, по Вурзил — 40%, по данным Луке —30%, по Дрейеру — 30—45%.
В практике кормления карпа определение количества белка в корме, которое обеспечивает быстрый рост рыб, методически более сложно, так как в силу особенностей питания оно в большей степени зависит от условий выращивания и технологии кормления, чем у форели.
В прудах, где потребности рыб в незаменимых факторах питания могут в определенной степени удовлетворяться за счет естественной пищи, содержание белка в корме может колебаться от 13—15% (зерновые) до 30—45% (комбикорма) и определяется комплексом факторов: естественной продуктивнетью водоема, плотностью посадки рыбы на единицуплощади, гидрохимическим и гидрологическим режимом, технологией приготовления кормов и техникой их скармливания.
При индустриальных методах выращивания карпа, предусматривающих исключительное питание рыб искусственными кормами, карп, подобно радужной форели, нуждается в более высоком содержании белка (в основном животного происхождения), что проявляется сильнее при неблагоприятных условиях среды.
По данным Акадзуки, полученным па синтетических диетах, для мальков карпа в возрасте до 3 месяцев оптимальным является содержание белка 40—45%. При количестве белка в рационе свыше 45% повышается смертность, при содержании ниже 20% наблюдается ухудшение роста.
Эксперименты Дренер, выполненные в аквариумах, показали, что для сеголетков карпа лучшие результаты дали форелевые корма с 30% белка, а для двухлетков и трехлетков — специальные карповые корма с 20—25% протеина.
Для канального сома потребность в белке в зависимости от различных факторов (при выращивании в прудах) колеблется в пределах 28—40%.
Для угря показана нецелесообразность повышения белка в корме свыше 40%.
Потребность в углеводах
Углеводы являются одним из главных источников обеспечения организма энергией. Их питательная ценность тесно связана с химическим строением и соотношением отдельных групп углеводов в кормах, а также со способностью пищеварительной системы рыб осуществлять расщепление и всасывание этих соединений.
По данным Филлипс, содержание переваримых углеводов в рационах форели не должно превышать 9—12% с учетом коэффициента их переваримости, принимаемого за 40%, т. е. 23—30% в сухом веществе диеты. Отмечено, что увеличение количества углеводов в корме приводит к патологическому накоплению гликогена в печени и повышенной смертности.
В то же время при соответствующей добавке витаминов высокое (порядка 50%) содержание углеводов в корме форели не ведет к перерождению печени и торможению роста.
Необходимость введения повышенных доз витаминов в высокоуглеводные корма для форели подтверждается исследованиями Шпанхофа. Его опыты на угре показали, что этот вид обладает способностью к более полному использованию углеводов корма, чем форель. При увеличении содержания крахмала до 50% изменений в скорости роста не наблюдали.
Карп способен утилизировать значительное количество углеводов. При выращивании его в прудах общее содержание углеводистых веществ может составлять около 70%.
Для нормального осуществления пищеварительных процессов рыбы должны получать с пищей достаточное количество трудногидролизуемых углеводов в качестве балластных веществ, придающих объемность концентрированным кормам, усиливающим перистальтику кишечника и частично активизирующим переваривание основных питательных веществ.
Потребность в жире
Жиры в кормах играют не только первостепенную роль в энергетическом обмене рыб, но являются источником незаменимых жирных кислот, фосфатидов и других соединений, способствующих лучшему усвоению питательных веществ корма и жирорастворимых витаминов.
При окислении они освобождают в два раза больше энергии, чем белки, и в несколько раз больше, чем углеводы корма.
С понижением температуры происходит перестройка жирнокислотного состава фосфолипидов в сторону увеличения ненасыщенности, что повышает текучесть жиров в условиях низких температур. С повышением температуры — наблюдается обратный процесс. Входя в состав липопротеинов — транспортной формы жиров — фосфолипиды принимают непосредственное участие в передвижении липидов в организме рыб, в том числе из кишечника, из печени и т.д. Триацилглицерины и фосфолипиды постоянно синтезируются в клетках из образующихся там и поступающих в процессе питания веществ, среди которых к незаменимым (эссенциальным) относятся высоконепредельные (полиненасыщенные) жирные кислоты. Они не синтезируются в организме и должны поступать с пищей в соответствии с потребностями рыб.
Поскольку незаменимые жирные кислоты составляют малую величину и, кроме того, эффективно удерживаются в организме, а источником энергии помимо жиров могут быть белки и углеводы, потребность рыб в жирах как поставщиках энергии не имеет четких границ. Маккей и Мацуи показали, что рыбы переносят значительные количества жира в пище. Лососи могут потреблять корма, содержащие до 57 % доброкачественного жира, а карп без видимых вредных последствий — около 40%. При дальнейшем повышении жира в рационе карпа до 80% рост рыб прекращался.
Точка зрения на необходимое содержание жира в рационе рыб неоднократно менялась. По данным Хигаши, мальки радужной форели могут болеть из-за отсутствия жира в пище, а взрослая рыба эффективно использует значительное количество жира. При этом для годовиков считалось оптимальным содержание жира до 5%, а для двухлетков до 15%.
Однако жиры обладают способностью к образованию токсичных продуктов окисления, вредное влияние которых на рыб установлено давно. Они вызывают торможение роста, анемию, жировое перерождение печени и почек, перерождение мускулатуры, а также могут являться канцерогенными веществами.
Несмотря на то, что была установлена белоксберегающая роль жира в рационах рыб, в 50-60-е гг. его предлагалось ограничивать в составе пастообразных лососевых кормов до 3-5% (Phillips, Podoliak, 1957; Rasmussen, 1961; Phillips, 1970 и др.). Это объяснялось широко распространенным в те годы заболеванием форели, потребляющей искусственные корма, жировой дегенерацией печени, которую часто связывали с обилием липидов в пище. Вместе с тем причиной этого заболевания, как выяснилось, было употребление недоброкачественных кормов с окисленным жиром. У форели, потреблявшей такие корма, развивалось цероидное перерождение печени, которое сопровождалось анемией и массовыми отходами рыб (Mann, 1952; Факторович, 1956, 1963; Остроумова, 1957; Хасимото, Окаити, 1968 и др.). Характерной особенностью заболевания было увеличение размера печени и ее песочная окраска.
Исследованиями, выполненными в 60-е гг. в ГосНИОРХ под руководством проф. Т.И. Привольнева (Привольнее и др., 1964, 1969), удалось установить, что включение в пастообразный рацион форели 10% подсолнечных фосфатидов предохраняет рыб от жировой дегенерации печени и сопутствующей ей анемии. При этом рыба лучше росла, имела минимальный отход, количество жира в печени снижалось более чем в 2 раза при повышении общей жирности рыб, содержание гемоглобина и эритроцитов нормализовалось. Далее было показано (Шестакова, 19766), что присутствие фосфатидов в корме обогащает печень форели фосфолипидами, особенно за счет лецитиновой фракции (фосфатидилхолин), отличающейся своими сильными антиокси-дантными свойствами. В зависимости от уровня фосфатидов в корме содержание фосфолипидов в печени увеличивалось следующим образом (Серпунина-Шестакова, 1979):
При определении потребности рыб в жирах имеет значение их качественный состав и количественное соотношение различных фракций липидов в животных и растительных жирах. У рыб, в частности у форелей, как и у теплокровных позвоночных, обнаружены симптомы недостаточности жирных кислот, что является признаком незаменимости их в питании.
ПОТРЕБНОСТЬ В УГЛЕВОДАХ
Углеводы — лабильные вещества, главный источник энергии в кормах теплокровных позвоночных. Основная масса углеводов содержится в растительных компонентах, в сухом веществе которых они могут достигать 80% и выше. В животном организме углеводы резервируются в небольших (несколько процентов сухого вещества) количествах в виде гликогена, преимущественно в печени и мышцах.
Природная пища рыб не богата углеводами, и большинство видов рыб не приспособлены к высокому содержанию их в рационе. Энергетические потребности покрываются у них в основном за счет белка и липидов. Большую роль у рыб играет процесс глюконеогенеза — образование глюкозы из неуглеводных предшественников (особенно из аминокислот), который в той или иной мере происходит у всех животных, но у низших, в том числе у беспозвоночных и рыб, его удельный вес выше, чем у более высокоорганизованных организмов.
Определенное количество углеводов пищи рыбы могут утилизировать, но эта способность неодинакова у разных видов: у теплолюбивых она выше, чем у холодолюбивых.
При разработке искусственных кормов стремятся снизить до минимума нерациональное использование белка в качестве источника энергии, сохраняя его для роста. При этом часть высокобелковых компонентов заменяют более дешевыми растительными.
Основной вопрос состоит в том, в каком количестве, при каких условиях и какие рыбы способны утилизировать растительные компоненты без потери роста и здоровья, почему эта способность различна у разных видов рыб.
Степень переваримости углеводов у рыб зависит от сложности их строения, от молекулярной массы. Простые сахара — моносахариды (глюкоза, галактоза и др.) — всасываются почти полностью у радужной форели, гольца, карпа, белого осетра и других видов рыб (Phillips, 1970; Щербина, 1973; Herold, Hung, 1995; Щербина, Гамыгин, 2006 и др.). Дисахариды, которые сначала подвергаются расщеплению, всасываются менее интенсивно, кроме мальтозы. Ее переваримость, например, у форели и осетра, одинакова с моносахаридами (свыше 99%). Дисахариды — сахароза — перевариваются в среднем и пределах 52-73%, лактоза у форели — 60, у осетра — 36%. Слабее сахаров расщепляются и всасываются полисахариды. Переваримость крахмала выше у карпа -19-48% к рациону.
В практике кормления углеводы делят на две большие группы: клетчатка и безазотистые экстрактивные вещества.
Клетчатка представляет собой основу оболочек растительных клеток, выполняет защитную и опорную функцию. Она состоит преимущественно из жестких полисахаридов — целлюлозы. Клетчатка относится к трудногидролизуемым углеводам, которые в последнее время стали называть «пищевыми волокнами». Помимо целлюлозы к ним относятся гемицеллюлоза, лигнин, пектин, хитин и др. (Щербина, Гамыгин, 2006).
В практике кормления животных клетчатку часто отождествляют с пищевыми волокнами. Подавляющее большинство позвоночных не имеет собственных ферментов, расщепляющих жесткие структуры клетчатки. Ее частичное переваривание осуществляется ферментами микрофлоры, населяющей пищеварительный тракт.
К безазотистым экстрактивным веществам относятся легкогидролизуемые углеводы, содержащиеся преимущественно внутри растительной клетки, — крахмал (главный резервный полисахарид растений) и различные сахара.
Эффективность использования растительных ингредиентов у рыб ограничивается двумя факторами — сравнительно низкой их переваримостью и проблемой утилизации избытка углеводов в обмене.
Потребность в минеральных элементАХ
Минеральное питание и потребности рыб в отдельных макро- и микроэлементах мало исследованы и в настоящее время являются предметом интенсивного изучения.
Трудности работ в этом направлении в методическом плане связаны с наличием осмотического пути проникновения элементов в организм рыб и со сложностью взаимодействия минеральных элементов в процессах обмена между собой, которое сопровождается частым возникновением антагонизма между отдельными ионами при нарушении определенных соотношений.
Известно, что потребность рыб в кальции может удовлетворяться почти полностью осмотическим путем, а в фосфоре за счет солей пищи. Изучение влияния концентрации кальция в воде и корме в условиях различного соотношения и концентраций кальция и магния в воде на рост двухлетних карпов показало, что увеличение карбонатного кальция в корме в два раза приводит к снижению его доступности организму рыб. На этот процесс оказывает действие не только абсолютное содержание кальция и магния в воде, но и их соотношение.
Известно, что на потребность рыб в минеральных элементах определенное влияние оказывает степень обеспеченности рациона витамином Д, а также форма, в которой минералы присутствуют в корме. Так, фосфор многих злаковых образует с инозитолом нерастворимые соли фитиновой кислоты (фитаты), имеющие прочную связь с кальцием. Эти соединения плохо доступны не только для теплокровных позвоночных, но также для лососевых и карповых рыб.
Установлено, что в минеральном обмене костистых рыб имеются видовые отличия, которые обусловливают различия в потребностях отдельных элементов. Исследования Огино свидетельствуют о том, что у форели полное отсутствие минеральных солей в корме вызывает через 2 недели замедление роста и повышенную смертность уже при проявлении первых симптомов недостаточности (отсутствие аппетита и вялость). В то же время у карпа в аналогичных условиях явных симптомов нарушения минерального обмена не наблюдалось. Отмечено лишь небольшое снижение темпа весового роста.
Определенное внимание уделяется и вопросам потребности различных объектов рыбоводства в микроэлементах, так как большая часть из них входит в состав групп ферментов и участвует в регуляции обмена веществ.
Качественные и количественные потребности рыб в макро- и микроэлементах зависят от следующих факторов:
· концентрации и соотношения солей в воде,
· качественного состава ингредиентов рациона,
· формы солей, в которых они присутствуют в кормах,
· их доступности организму рыбы
· степени обеспеченности рациона другими питательными веществами, необходимыми для нормального течения метаболизма.
Потребность в витаминах
Витамины представляют собой низкомолекулярные биологически активные органические соединения, которые в очень малых количествах абсолютно необходимы для жизнедеятельности организма. Витамины участвуют в обмене веществ, играя роль стимуляторов и регуляторов физиолого-биохимических процессов. Входя в коферменты и простетические группы белков, они выступают как постоянные биологические катализаторы химических реакций. От присутствия витаминов зависят нормальное развитие и рост рыб, воспроизводительная функция, устойчивость к стрессам и болезням.
По растворимости в разных средах витамины делятся на две группы: жирорастворимые (витамины А, Е, D, К) и водорастворимые (витамины С и группы В).
Большинство витаминов не синтезируются в организме рыб и должны поступать с пищей. Естественная пища рыб богата витаминами. У наземных жвачных животных источником витаминов группы В и К служит микрофлора пищеварительного тракта. Среди рыб микробный синтез витаминов играет особенно заметную роль у видов, обитающих в условиях высоких температур и имеющих сравнительно длинный объемистый кишечник (растительноядные). У рыб витамин А поступает как в готовом виде с животной пищей, так и производится из предшественников — каротиноидов, получаемых с кормом. Другие же, в том числе лососевые, сиговые, осетровые, если и обладают этой способностью, то не в такой степени, чтобы покрыть потребности. Именно поэтому форель, в отличие от карпа, очень чувствительна к дефициту аскорбиновой кислоты в корме.
Витамин С некоторые рыбы, например карпы, караси, способны синтезировать. Икеда и Сато считают, что карп может синтезировать определенное количество аскорбиновой кислоты, но оно недостаточно для обеспечения нормального роста молоди. По мнению Хасимото, потребность карпа в витамине B12 обеспечивается при помощи его кишечной микрофлоры.
Благодаря серии исследований американских и японских ученых на экспериментальных диетах была установлена потребность в водорастворимых витаминах для таких объектов холодноводного рыбоводства, как радужная форель, чавыча, кижуч, нерка, сима, голец, кумжа. При этом было показано, что специфические синдромы недостаточности витаминов одинаковы для большинства видов рыб. Признаки гиповитаминозов изложены в сводной таблице, составленной И. Н. Остроумовой (1974а) по данным Филлипс (Phillips, 1970) и Хальвера (Halver, 1972).
В общем виде в начале 80-х лет потребность форели в водорастворимых витаминах выражалась в следующих величинах (мг/кг веса рыбы в сутки):
· тиамин (B1) 0,13—2,0;
· рибофлавин (В2) 0,75—1,0;
· пиридоксин (В6) 0,38—0,43;
· пантотеновая кислота (В3) 1,3—2,0;
· холин (В4) 50—70;
· никотиновая кислота (В5, РР) 5—7;
· цианкобламин (В12) 0,0002—0,0003;
· фолиевая кислота (Во), 0,10—0,15;
· биотин (Н) 0,03—0,04;
· инозитол 18—20;
· аскорбиновая кислота (С) 2—3.
Величина потребностей в витаминах у рыб и симптомы их недостаточности являются предметом пристального изучения на протяжении всего активного периода развития интенсивных форм рыбоводства, т.е. начиная с 50-60-х гг. Вплоть до последнего времени они постоянно рассматриваются и корректируются. Обращают на себя внимание значительно более низкие уровни потребностей для форели по данным NRC, 1993 (Национальный научный совет США), особенно среди водорастворимых витаминов группы В, по сравнению со сведениями, приведенными в обобщениях начала 80-х гг. Хальвером и Штеффенсом (Halver, 1982; Steffens, 1985), и последующими данными.
Сложность изучения потребностей в витаминах у рыб состоит в том, что они зависят от многих факторов — вида и возраста рыб, этапа жизненного цикла (личинки, молодь, производители в период созревания гонад и нереста и т.д.), физиологического состояния, стресса, температуры воды, содержания витаминов в кормовых компонентах,
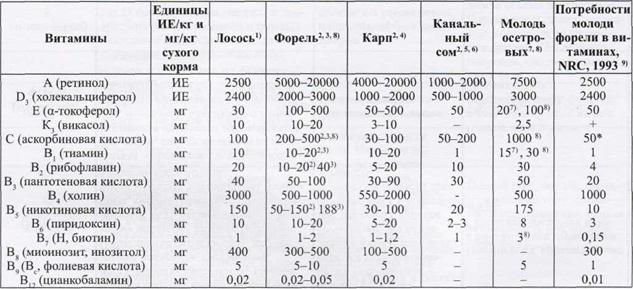
Количественные потребности в различных витаминах имеют видовые отличия даже среди лососевых (Табл.1).
При отсутствии того или иного витамина в корме у рыб возникают авитаминозы; при недостаточном поступлении, что в практике встречается чаще, — гиповитаминозы.
Длительное поступление значительных (во много раз превышающих норму) количеств витаминов приводит к гипервитаминозам. Последние случаи отмечены в основном для жирорастворимых витаминов, которые удерживаются в организме более продолжительное время, чем быстро вовлекаемые в обмен и выводимые водорастворимые.
Симптомы отсутствия или недостатка витаминов у тепловодных видов рыб аналогичны таковым у холодноводных видов. Как показали исследования, в количественном отношении карп менее чувствителен к недостатку витаминов, и симптомы гиповитаминоза у него проявляются значительно позднее, чем у форели и других лососевых.
О различиях в потребности в витаминах у отдельных видов рыб свидетельствуют также данные Л. Шпанхофа, согласно которым речной угорь менее чувствителен к недостатку витамина В4, чем радужная форель.
Известно, что при недостатке белка в пище тормозится усвоение некоторых витаминов (А, В2, В5). При повышении уровня белка увеличивается потребность в витаминах В1; B6, С, а при избытке жира — в витамине Е. Обмен аминокислот тесно связан с обменом отдельных витаминов. При недостатке метионина повышается потребность в витамине В12. При дефиците никотиновой кислоты она может синтезироваться из триптофана и тем самым снижать резервы этой незаменимой аминокислоты, что отражается на белковом росте (Таблица 1).
Недостаток жира в корме приводит к снижению всасывания и нарушает обмен жирорастворимых витаминов. С другой стороны, при высоком содержании жира дефицит витаминов-антиоксидантов (Е, А, С и др.) приводит к развитию перекисного свободнорадикального окисления липидов.
Таблица 1
Количественные потребности в витаминах у рыб
Отметим еще способность витаминов к взаимодействию и взаимозаменяемости. Наибольшей заменяющей потенцией, установленной на теплокровных животных, обладает аскорбиновая кислота (Терруан, 1969), которая иногда может полностью снять симптомы дефицита других витаминов. Удивительные свойства этого витамина хорошо известны и в практике форелеводства, а теперь уже и осетроводства.
В отношении жирорастворимых витаминов вопрос оказался более сложным. В результате большой серии экспериментов показано, что витамины А (ретинол), Д (кальциферол), Е (токоферол) и К (викасол) необходимы для поддержания функциональной активности и обеспечения процессов роста рыб так же, как и теплокровных животных.
Данные о количественных потребностях в витаминах этой группы менее определенны, и существующие разработки свидетельствуют, что при введении синтетических витаминов этой группы чаще возникает гипервитаминоз, который вызывает торможение роста рыб.
Проведенные исследования послужили основой для составления витаминных премиксов, которые применяются в нашей стране и за рубежом.
6.3. способы приготовления Искусственных кормов
Рыбоводные хозяйства нашей страны используют для выращивания рыбы стартовые и продукционные комбикорма, полученные путем:
1) прессования комбикормов, увлажненных сухим паром;
2) прессования комбикормов, увлажненных водой;
3) центробежного гранулирования;
4) микрокапсулирования (нанесение водозащитного покрытия на поверхность);
5) прессования предварительно экструдированных компонентов с последующим измельчением и увлажнением сухим паром;
6) экструдирования.
При изготовлении кормов связующими элементами являются также отдельные компоненты рыбных кормов, такие как пшеничная, водорослевая и кровяная мука, сухой обрат.
Стартовые комбикорма получают первыми четырьмя способами. Отечественные стартовые комбикорма: для лососевых
- РГМ-8М (для p.Salmo),
- ЛС-НТ (низкотемпературного оптимума действия для тихоокеанских лососей),
- МКС-1-86 и СГК-88 (для p. Oncorhynchus); для сиговых
- РГМ-СС (стартовый сиговый - для личинок и ранних мальков),
- РГМ-ПС (продукционный сиговый - для мальков и сеголетков),
- РГМ-ЛБ (для личинок белорыбицы), -ЛС-81,
- МС-84;
для осетровых -ОСТ-4, ОСТ-6.
Продукционные комбикорма получают всеми способами, кроме третьего и четвертого.
Влажное прессование комбикормов для рыб на 19-22 % снижает кормовые затраты по сравнению с комбикормами сухого прессования и на 28 % по сравнению с тестообразными комбикормами. По сравнению с последними рыбоводный эффект гранулированных комбикормов с водозащитным покрытием повышается на 15 %, гранулированных комбикормов из тонко измельченных компонентов - на 16,7, брикетированных - на 18 %.
Использование экструдированных (плавающих) кормов позволяет уменьшить кормовые расходы на единицу прироста рыбы до 1,3 - 1,5. Экструдаты в течение одних суток могут находиться на плаву. Экстракция питательных веществ из экструдатов вдвое меньше, чем из гранул.
Экструдированные комбикорма (экструдаты) производятся с помощью экструдера (например, отечественный КМЗ-2). В процессе экструдирования компоненты комбикорма под одновременным воздействием возрастающих значений температуры (80 - 100 - 120°С), влажности (17 - 20 - 25 %) и давления (0,56 - 0,32 МПа) приобретают свойства текучести и выпрессовываются под действием шнека через отверстия головки экструдера. Вследствие резкого перепада давления аккумулированная комбикормом энергия освобождается с мгновенной скоростью, в результате чего происходит эффект взрыва, приводящего к резкому выходу пара из структуры материала. Этот эффект вызывает образование пористой структуры экструдата с объемной массой 180 - 320 г/л.
Химический состав экструдатов такой же, как и гранулированных кормов. Экструдаты в зависимости от диаметра (в мм) делятся на следующие группы:
1-я - 1,7-2,5;
2-я - 2,5 - 3,7;
3-я - 3,7 - 5,0;
4-я - 5,0 - 7,0;
5-я - 7,0-9,0.
Экструдаты должны плавать на поверхности воды не менее 30 мин.
В настоящее время функционируют четыре специализированных завода по воспроизводству специальных комбикормов, предназначенных для выращивания рыбы в замкнутых системах, садках и бассейнах. Два из них в России (Ростовский на Дону и Белгородский), по одному на Украине (Днепропетровский) и в Узбекистане (Чиназский). Стартовые и продукционные комбикорма вырабатывают на всех заводах. На Днепропетровском предусмотрено производство экструдированных комбикормов, на Белгородском - микрокапсулированных.
Основная цель микрокапсулирования - предотвратить или свести до минимума потери питательных веществ из комбикорма в период нахождения его в воде. Водозащитное покрытие на поверхность стартовых комбикормов наносится во взвешенном состоянии, которое обеспечивается сжатым воздухом. В качестве пленкообразующих материалов используются связующие материалы (крахмал картофельный, гумат натрия, пектин) и прилипатели (стеариновая кислота, стеарин кальция, парафин, различные масла, изобутелен, желатин, клей и др.).
Эффективность кормления рыбы оценивают по кормовому коэффициенту, темпу роста, выживаемости, физиологическому состоянию рыбы, стоимости корма.
Кормовой коэффициент (КК) это количество корма, которое необходимо затратить, чтобы получить единицу весового прироста. Например, для молоди осетровых кормовой коэффициент дафний равен 6, олигохет - 2, артемии - 4; для лососевых КК дафний - 6-7, олигохет - 3-4. Другими словами, КК выражает отношение съеденного к приросту.
Различают также истинный кормовой коэффициент (ИКК) и рабочийкормовой коэффициент (РКК), или коэффициент оплаты корма (КОК).
ИКК - это отношение количества корма, непосредственно съеденного рыбой, к приросту ее массы. Определяют его экспериментальным путем, применяя физиологические методы или прямые наблюдения за потреблением корма.
РКК отражает отношение количества заданного корма к приросту рыбы. РКК всегда больше ИКК, так как часть корма теряется. Даже для одного и того же корма и одного и того же вида рыбы эти коэффициенты не остаются постоянными.
Затраты корма на единицу прироста зависят от качества корма, вида рыбы, ее возраста, физиологического состояния, температуры воды, газового режима, гидрохимических условий, техники кормления, освещенности. У молодых рыб кормовые коэффициенты всегда меньше, чем у более старших. У здоровой рыбы эти показатели меньше, чем у больной и ослабленной. Если корм задается рыбе несколько раз в сутки мелкими порциями, то показатели ИКК и РКК будут ниже, чем при одновременном скармливании больших количеств того же корма.
При кормлении рыб большое значение имеет суточная ритмика и нормы внесения корма в зависимости от возраста и условий выращивания. В частности, при увеличении индивидуальной массы рыбы относительная величина рациона уменьшается, а при повышении температуры воды до оптимума - возрастает. Серьезное внимание необходимо уделять способам внесения корма, механизации и автоматизации этого процесса.
Контрольные вопросы к главе 6
1. Как развивались представления о процессе питания?
2. Охарактеризуйте теорию сбалансированного питания.
3. Опишите теорию адекватного питания.
4. Что понимается под балластными веществами и как менялось представление об их роли в процессе питания?
5. Почему особь рыбы может рассматриваться как надорганизменная система?
6. Какова роль микрофлоры в процессе пищеварения рыб?
7. В каких случаях организм переходит к эндотрофии?
8. Опишите концепцию универсальных функциональных блоков в биосфере.
9. Охарактеризуйте химический состав естественной пищи рыб.
10. Что понимают под сбалансированным рационом рыб?
11. Охарактеризуйте потребности рыб в белках, жирах и углеводах.
12. У каких видов рыб отмечаются высокие потребности в белках?
13. Почему высокое потребление жира приводило к заболеваниям печени у форели в середине ХХ столетия?
14. Охарактеризуйте потребность рыб в витаминах.
15. Какие способы приготовления искусственных кормов Вы знаете?
16. Какие корма называются стартовыми?
17. Какие корма называются продукционными?
18. В чем разница экструдированных и микрокапсулированных кормов рыб?
19. Что понимают под кормовым коэффициентом и чем он отличается от рабочего кормового коэффиционта и коэффициента оплаты корма?
ГЛАВА 7
Важной биологической основой рыбоводства является способность рыб к росту, а у ряда видов – к высокой скорости массонакопления, что является основой производства ихтиомассы в рыбоводстве. Зависимость скорости роста от интенсивности питания имеет асимптотическую зависимость (рис. 1).
Логарифмическая фаза роста заканчивается перед первым нерестом, а пре-дельная масса рыб в 2,5 - 6 раз больше, чем при первом нересте (Яржомбек,1996).
Некоторые рыбы - дальневосточные лососи, голомянки, угри и др. - гибнут после первого нереста и, таким образом, как бы растут всю жизнь. Многие долгоживущие и испытывающие значительный пресс хищников и промысла рыбы практически никогда не доживают до предельного возраста и тоже как бы растут всю жизнь. Рациональное рыболовство и товарное рыбоводство основаны на эксплуатации достаточно молодых, все еще быстро растущих рыб.
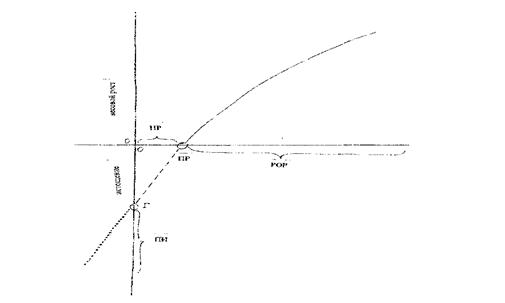
Рис.1. Изменения массы тела (рост и истощение): Г- голодание,
HP - недостаточный рацион, ПР - поддерживающий рацион, POP - рацион
обеспечивающий рост, ПИ — потери при икрометании, эякуляции,
кровопотерях, слизеотделении
Рост рыб может иметь место, только если рацион превышает потребности в пище, обеспечивающие покрытие расходов организма. Потенциальная способность к росту (максимальный возможный прирост) генетически обусловлен, то есть величине прироста определён некоторый предел: есть быстрорастущие рыбы и тугорослые. Никакими условиями питания невозможно заставить организм превысить потенциальную способность к росту. Максимальный прирост возможен при максимальной интенсивности питания в оптимальных условиях обитания. При понижении интенсивности питания рост замедляется.
При поддерживающем питании масса остаётся постоянной, при превышении поддерживающего рациона прирост сначала увеличивается пропорционально интенсивности питания (величине рациона), а затем увеличение рациона всё в меньшей степени ускоряет рост. При превышении некоторой величины рациона скорость роста перестаёт увеличиваться. Это происходит потому, что при дальнейшем увеличении рациона понижается усвоение пищи. Максимальное использование пищи на рост (30-50%) наблюдается при небольшом рационе - 20-30 % от максимального. При максимальном рационе на рост используется приблизительно 1/10 часть питательных веществ пищи.
Питание компенсирует затраты веществ, производящиеся при жизнедеятельности - энергетические траты при окислении веществ, при анаэробном распаде веществ, потери отработанных веществ - витаминов, нуклеотидов, низкомолекулярных органических соединений, минеральных компонентов тела - соединений макроэлементов и микроэлементов. При полном отсутствии питания (голодании) происходит потеря массы тела и истощение резервов. При недостаточном питании истощение происходит медленнее, чем при полном голодании, но также может наблюдаться потеря массы или замедленный рост. При неполноценном питании возникают болезненные явления, подобные авитаминозам.
Прирост массы тела рыбы - накопление его питательной части - белков, жиров, витаминов, минеральных биогенов связан с потреблением кормов и происходит "по остаточному принципу": поступление веществ в организм сначала должно компенсировать постоянно происходящие в живом организме потери, вызванные метаболизмом, и только превышение поступлений над потерями ведет к приросту. Для роста тела необходим положительный баланс потребляемых и расходуемых соединений. Этот процесс рассматривается как биологические ресурсы роста. Основную часть сухого вещества тела составляют органические вещества - протеин, жир, нуклеиновые кислоты, углеводы, общее количество которых может быть выражено в виде калорийного эквивалента, или эквивалентного количества, кислорода, необходимого для окисления. Это дало возможность Г.Г.Винбергу (1956) выразить основные постулаты баланса органики и энергии у рыб. В настоящий период достижения биохимии и физиологии рыб позволили по-новому раскрыть суммарные балансовые расчеты массонакопления. Органические вещества представляются в настоящее время в виде незаменимых и заменяемых, дефицитных и ограниченных к потреблению (Яржомбек,1996). Значительное освещение получил баланс минеральных биогенов, механизм формирования потребностей в них и пути удовлетворения потребностей. Выяснены принципы количественного учета в реальном балансе вещества и энергии расходов на двигательную активность рыб, на анаболические процессы, обусловленные питанием. Для количественных расчетов достаточно известно о количественном выражении пищеварительных процессов, биосорбции из воды растворенных веществ.
При изучении процесса прироста массы важно знание усвоения веществ при кормлении. Абсолютным показателем является количество вещества, усваиваемое в единицу времени в расчете на единицу массы тела (мг/кг массы тела). Относительным показателем скорости усвоения является показатель доли вещества, усваивающегося в единицу времени, или время усвоения определенной части вещества, например время усвоения половины вещества. Итоговым показателем является полнота усвоения.
Для изучения пищеварительных процессов (Рис.2) применительно к своим интересам и возможностям после ряда попыток в разных направлениях был разработан метод принудительного кормления карпа с последующим промыванием кишечника (Яржомбек, Щербина, Здор, Бекина, 1980).
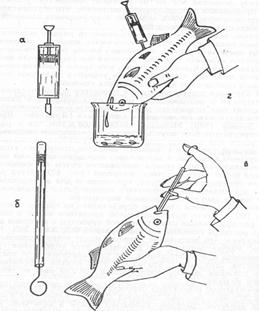
|
Рис. 2. Приемы изучения пищеварения карпа с принудительным кормлением:
а - щприц с мягкой насадкой для промывания кищечника; б - трубка с поршнем для введения вещества в кишечник; в - введение вещества в кишечник; г - промывание кишечника и получение его содержимого
Количественные показатели ассимиляции и диссимиляции ве¬щества являются основой для научного подхода к решению задач рационального рыбного хозяйства. Ассимиляция как приходная часть баланса включает в себя питание, потребление кислорода и биосорбцию веществ из воды.
Диссимиляция как расходная часть баланса - это траты вещества в катаболических процессах, активная экскреция, пассивная потеря веществ и выделение половых продуктов. Эти явления определяют эффективность питания, обеспеченность ресурсами в естественной среде и в условиях рыбоводного хозяйства. Кроме того, знание закономерностей абсорбции, усвоения и экскреции веществ весьма важно для токсикологии - одной из главных проблем настоящего и будущего обитателей гидросферы. Сложность балансовых процессов обусловлена, с одной стороны, многокомпонентностью ресурсов роста, а с другой - множественностью влияний внешних и внутренних факторов: это видовые особенности рыб, влияние размеров тела, температуры, мышечной работы, питания и др. (Рис.3).
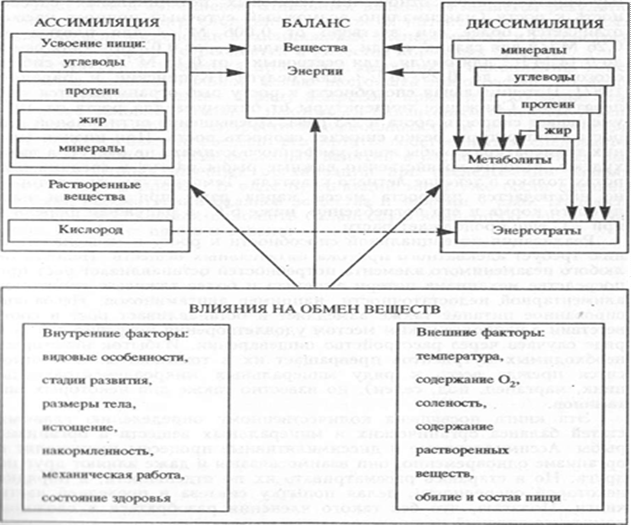
Рис.1. Схема баланса процесса массонакопления у рыб.
Отрицательный баланс возникает в случае превышения диссимиляции над ассимиляцией во время голодовок, зимовок, миграций и нереста. Однако отрицательный баланс одних веществ может сопровождаться положительным балансом других. И наоборот - питание и рост могут сопровождаться постепенным истощением запасов какого-либо важного вещества, пока его недостаток не станет патологическим.
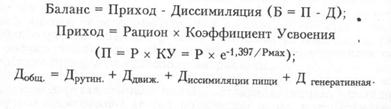
Потенциальная способность к росту у рыб до полового созревания может быть выражена в виде обратной пропорциональности от кубического корня массы тела, что является отражением так называемого закона поверхности. Максимальный суточный прирост является видовым признаком. В разных систематических группах рыб имеются быстро- и медленнорастущие виды. Например, среди изученных в этом отношении карповых потенциальная способность к росту (максимально возможный суточный прирост массы) отличается более чем вчетверо - от 0,006 М-1/3 для плотвы до 0,26 М-1/3 для сазана, а среди лососевидных - от 0,07 М-1/3 для семги до 0,14 М-1/3 для омуля, для осетровых - от 0,13 М-1/3 для сибирского осетра до 0,29 М-1/3 для белуги (Купинский, Баранов, 1987).
Потенциальная способность к росту рыб ограничивается температурой. Снижение температуры от оптимума для роста на 10°С уменьшает скорость роста в 2-3 раза. Превышение оптимальной для роста температуры резко снижает скорость роста. При низких зимних температурах рыбы зоны умеренного климата не растут, а даже худеют. У карпа при температуре 6°С и ниже не наблюдается прироста массы даже при наличии лучших кормов, а радужная форель растет и при 2°С.
Реализация потенциальной способности к росту и массонакоплению требует адекватного притока питательных веществ. Недостаток любого незаменимого элемента потребностей останавливает рост при посредстве механизма потери аппетита и более тяжелых симптомов алиментарной недостаточности, например, авитаминозов. Несбалансированное питание также замедляет и останавливает рост в соответствии с наиболее узким местом удовлетворения потребностей, а в ряде случаев через расстройство пищеварения. Избыток некоторых необходимых нутриентов превращает их в токсиканты. Это относится, прежде всего, к ряду минеральных микроэлементов: медь, цинк, марганец, йод, селен, но известно также для некоторых витаминов.
Главнейшими рыбоводными задачами является количественное определение главных компонент баланса органических и минеральных веществ в организме рыбы. Ассимилятивные и диссимилятивные процессы происходят в организме одновременно, они взаимосвязаны и даже влияют друг на друга. Расходная часть баланса представляет уровень трат который зависит от деятельности различных систем - двигательной, метаболической, генеративной. Приходная часть баланса определяется питанием, пищеварением, всасыванием, а также биосорбцией через внешние структуры.
Биосорбция в отношении веществ, кроме молекулярного кислорода, является особенностью гидробионтов и мало известна общей физиологии.
Для технологии товарного рыбоводства основным является свойство рыб к накоплению массы тела. Практически весь технологический процесс выращивания ихтиомассы построен на управлении скоростью массонакопления. При этом используются зависимости накопления массы от начальной массы рыб, генетических особенностей вида и экологических условий.
Зависимость скорости массонакопления от начальной массы рыб проявляется в самозамедлении роста: чем выше индивидуальная масса рыбы, тем больше время удвоения этой массы. Эта закономерность нашла отражение в нормативной документации по выращиванию карпа. Так, личинки карпа удваивают массу в среднем ежесуточно, мальки – каждые 4-5 суток, сеголетки – каждые 10-15 суток, двухлетки – каждые 20-30 суток.
Под влиянием генетических особенностей на скорость роста понимают в рыбоводстве не только быстрый рост или тугорослость, но и различия внутри видовых форм, например, связанных со сроками и местами размножения или типами питания, а иногда и теплолюбивостью, закрепленной генетически у породы, как это наблюдается у украинских пород карпа.
Влияют на скорость роста и экологические факторы среды, которые чем полнее удовлетворяют оптимуму вида, тем полнее могут обеспечить максимальные весовые приросты рыб. В связи с этим любой фактор среды может быть оценен по его продуктивному действию, то есть по способности влиять на результат рыбоводного процесса. Так, температуру в диапазоне 25-30°С для роста карпа считают оптимальной, а 36°С – пороговой.
Для характеристики каждого фактора среды рыбоводы пользуются двумя шкалами – качественной экологический и количественной физической. Качественная экологическая шкала включает такие оценки благоприятные, неблагоприятные, оптимальные, субоптимальные, запороговые, подпороговые, пороговые условия. При этом каждый параметр среды может быть охарактерихован величиной, так температура – в °С, содержание кислорода в воде и других химических веществ в мг/л, мл/л, концентрация организмов пищи в естественной кормовой базе в г/м2, г/м3 и т.д.
Классическим примером влияния факторов на рост рыб является резкое изменение темпа роста атлантического лосося, или семги в связи с изменением условий обитения. В первые годы жизни семги в реке при питании личинками насекомых семга растет очень медленно. Скатившись в море в 3-4 летнем возрасте и перейдя на питание рыбой ее скорость роста значительно возрастает: если к 4 году средняя масса достигала 1370 г (по данным Берга), то к 8 годам – 9150 г.
Контрольные вопросы к главе 7
1. Нарисуйте кривую роста массы рыб и выделите на ней периоды голодания и интенсивного роста.
2. Как связаны весовой рост рыб и питание.
3. В каких исследованиях используют метод принудительного кормления карпа?
4. Какие экологические факторы влияют на рострыб?
5. Как связан рост рыб с генетическими факторами?
6. В каком диапазоне изменяется величина максимально возможного суточного прироста массы у карповых рыб?
7. В каком диапазоне изменяется величина максимально возможного суточного прироста массы у лососевидных рыб?
8. В каком диапазоне изменяется величина максимально возможного суточного прироста массы у осетровых рыб?
9. Как зависит скорость массонакопления от температуры?
ГЛАВА 8
|
|
|
Дата добавления: 2014-01-06; Просмотров: 3496; Нарушение авторских прав?; Мы поможем в написании вашей работы!