КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Мини-сайзер с двумя матрицами
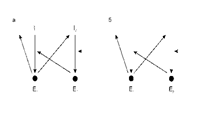
• а – сайзер с несцепленными матрицами,
• б – сайзер со сцепленными матрицами.
МИНИ-САЙЗЕР С ДВУМЯ СЦЕПЛЕННЫМИ МАТРИЦАМИ
• 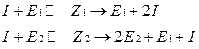 Схема кинетических процессов в данном случае будет иметь вид:
Схема кинетических процессов в данном случае будет иметь вид:
Эта система является простейшей и допускает более подробное исследование.
Обозначим полные концентрации фракций I, Е1 и Е2 через x1, x2 и x3 соответственно.
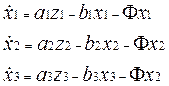 Уравнение динамики имеет стандартный вид:
Уравнение динамики имеет стандартный вид:
где z3=z2 и введено только для симметричности записи,
а условия квазиравновесия комплексов имеют вид
k1z1 = (х1 — z1)(x2 — z1),
k2z2 = (х1 — z2)(x3 — z2),
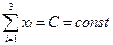 |
при селекционном ограничении
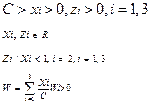 и дополнительных ограничениях:
и дополнительных ограничениях:
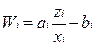 |
где
«селективные ценности» классов макромолекул
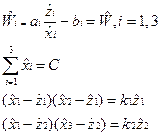 Условия нетривиального равновесия, когда сайзер существует
Условия нетривиального равновесия, когда сайзер существует
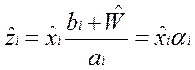 Используя уравнение на селективные ценности, выразим концентрации комплексов через полные концентрации:
Используя уравнение на селективные ценности, выразим концентрации комплексов через полные концентрации:
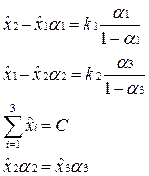 Подставив эти выражения в уравнения на комплексы и исключив переменные Zi,получим систему равенств на равновесные концентрации и селективные ценности.
Подставив эти выражения в уравнения на комплексы и исключив переменные Zi,получим систему равенств на равновесные концентрации и селективные ценности.
Решая эти уравнения относительно xi, получаем равновесные концентрации
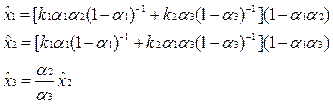 |
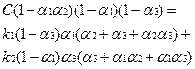 Суммируем стационарные значения xi, пользуемся тем, что сумма должна быть равна С и после некоторых преобразований получаем
Суммируем стационарные значения xi, пользуемся тем, что сумма должна быть равна С и после некоторых преобразований получаем
• 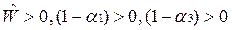 При этом из тех условий, что были выписаны ранее, вытекает, что
При этом из тех условий, что были выписаны ранее, вытекает, что
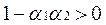 Поэтому для выполнения равенства (***) необходимо, чтобы
Поэтому для выполнения равенства (***) необходимо, чтобы
•
• 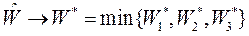 Кроме того, из уравнения (***) видно, что при стремлении средней стационарной селективной ценности к верхним значениям
Кроме того, из уравнения (***) видно, что при стремлении средней стационарной селективной ценности к верхним значениям
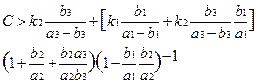 Левая часть равенства (***) стремится к 0, а правая остается конечной. Тогда можно найти достаточное условие существования нетривиальной равновесной точки
Левая часть равенства (***) стремится к 0, а правая остается конечной. Тогда можно найти достаточное условие существования нетривиальной равновесной точки
• 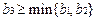 Достаточным условием локальной устойчивости этой точки покоя оказывается неравенство
Достаточным условием локальной устойчивости этой точки покоя оказывается неравенство
Таким образом, существование и локальная устойчивость нетривиальной равновесной точки обеспечены при довольно слабых ограничениях на параметры.
У системы есть еще 4 тривиальные и полутривиальные равновесные точки. Иследование локальной устойчивости этих точек показывает, что достаточные условия существования и локальной устойчивости нетривиальной равновесной точки достаточны также для неустойчивости всех остальных точек покоя системы.
• Таким образом, достаточные условия, приведенные выше, обеспечивают невырождение сайзера со сцепленными матрицами и локальную устойчивость нетривиальной равновесной точки, хотя и не исключают существования более сложных режимов, в том числе периодических.

Как и в задаче о сайзере с несцепленными матрицами, биологический смысл этих условий существования и устойчивого функционирования сайзера заключается в:
• достаточно высокой плотности вещества и
• достаточно высокой скоростью распада белка трансляции.
Однако, ограничения на параметры более слабые, чем в случае несцепленных матриц, следовательно, область допустимых параметров более широкая, а система в целом более легко реализуемая.
Мини-сайзер с двумя сцепленными матрицами
• Нелинейность системы и зависимость приспособленностей фракций от концентраций приводят к тому, что средняя приспособленность коллектива макромолекул достигает максимума не в устойчивой равновесной точке, а в другом месте, т. е. фундаментальная теорема естественного отбора Фишера не выполняется. Эта теорема была доказана Рональдом Фишером (Ronald Fisher), и она гласит: «Скорость повышения приспособленности в любой популяции в любой момент времени равна ее генетической дисперсии по приспособленности в этот момент». Теорема имеет двоякий смысл: она формально доказывает невозможность отбора в генетически однородной популяции; она показывает, что скорость изменения приспособленности определяется не ее значением, которое создает конкретный аллель, а степенью изменчивости по приспособленности, создаваемой в популяции всеми аллелями данного гена.
• Таким образом, в целом, сцепление матриц играет стабилизирующую роль в динамике сайзеров.
Закономерности эволюции сайзеров
• Можно провести исследования способности сайзера к эволюции путем мутаций по параметрам модели. Предположим, что исходная матрица (I1) порождает мутантную (I2), кодирующую те же белки, но имеющую другие параметры. Тогда, используя Эйгеновский подход, можно показать аналитически, что сайзер со сцепленными матрицами становится структурно неустойчивым уже при добавлении одной мутантной фракции. По мутациям, затрагивающим приспособленность сайзера (а это в первую очередь кинетические параметры), происходит типичный дарвиновский движущий или стабилизирующий отбор. Вполне вероятно образование «шлейфа» паразитических матриц, имеющих такие же параметры воспроизведения, но не производящих белки.
• Внутренней оптимизации по концентрациям матриц внутри сайзера не происходит. Тем не менее, равновесные сайзеры имеют разные средние приспособленности, т. е. разные темпы роста и возможности для взаимной конкуренции. Конкуренция двух сайзеров в одном «ящике Эйгена» невозможна из-за универсальности процессов. Возможна, однако конкуренция сайзеров, уравновешенных в разных «ящиках», в общем метаболическом потоке. Изменение пропорций концентраций матриц происходит случайным образом, но закрепляется в итоге конкуренции.
• Поскольку сайзеры не имеют ограничений на число генов внутри единой матрицы, а также на число кодируемых белков и на сложность их функциональных отношений в системе, то именно таким путем может происходить усложнение сайзеров. Увеличение числа типов матриц в этом случае не нужно.
• Следует отметить, что при эволюционном переходе к матрицам, которые содержат более одного гена, требуется дискретность этих генов. Т.е., становится необходима трансляционная пунктуация, которая может иметь очень раннее эволюционное происхождение.
• Итак, сайзеры обладают широким спектром динамических свойств:
• 1) способностью к внутреннему уравновешиванию и невырождению,
• 2) к росту,
• 3) к внутренней эволюции путем отбора мутантных вариантов матриц,
• 4) к конкуренции.
• При этом устойчивый полиморфизм мутационных вариантов почти невозможен, в виду того, что сайзеры находятся вблизи границы структурной устойчивости.
|
Дата добавления: 2014-01-07; Просмотров: 413; Нарушение авторских прав?; Мы поможем в написании вашей работы!