КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Квазивиды. Катастрофа мутационных ошибок
|
|
|
|
•  Динамику комплекса самовоспроизводящихся молекул можно описать следующей системой дифференциальных уравнений (Эйген, Шустер, 1982)
Динамику комплекса самовоспроизводящихся молекул можно описать следующей системой дифференциальных уравнений (Эйген, Шустер, 1982)
• где i — индекс различных типов самовоспроизводящихся молекул, характеризующий закодированную в них информацию. Символ Xi обозначает i-ю популяционную переменную или ее концентрацию. Данная система уравнений включает в себя те самовоспроизводящиеся единицы, которые присутствуют в рассматриваемой системе и которые могут быть пронумерованы от 1 до N. Если в ходе эволюции появляются различные мутанты, то система может быть расширена.
• Эти уравнения описывают открытую систему. В данной формализации метаболизм представлен спонтанным синтезом (AiQixi) и дезинтеграцией (Dixi) молекулярного вида. «Спонтанность» в данном контексте означает, что обе реакции не являются взаимно обратимыми. Член Ai содержит какую-либо стехиометрическую функцию fi(m1, m2..... mλ) концентраций высокоэнергетического строительного материала, необходимого для синтеза молекулярного вида i. Точная форма функции зависит от конкретных механизмов реакций. Этот высокоэнергетический строительный материал постоянно поступает с притоком вещества, а продукты реакций элиминируются соответствующим оттоком (Фi). Член Di, описывает спонтанное разложение. Он линейно связан с xi по обычному закону первого порядка. Однако, в более сложных системах как синтез, так и деградация могут катализироваться ферментами, реакции могут иметь дополнительные связи, и тогда как Ai, так и Di могут включать в себя другие функции концентраций.
• Самовоспроизведение — описывает образование xi как функцию от xi. Простейшая форма автокатализа описывается прямой линейной зависимостью. Аналогичным образом можно рассматривать более сложные линейные механизмы, например комплементарное инструктирование (синтез матрицы по матрице) или циклический катализ.
• Мутабильность описывается через фактор качества Qi, который может принимать любое значение между нулем и единицей. Значение Qi определяет ту долю репродуктивных копий, которые образуются на данной матрице с образованием ее точной копии. Дополнительный член — Ai (1 - Qixi), относится к неточной репродукции матрицы i. Он описывает образование разнообразных «ошибочных копий». В большинстве случаев эти копии очень мало отличаются от вида i.
• Появление этих ошибочных копий вида i учитывается соответствующими членами в динамических уравнениях для его «родственников» k. А копия i получает вклады от этих «родственных видов» из-за ошибок в их репликации. Они учитываются суммой. Значение параметра wki, задающего индивидуальный темп мутаций, тем меньше, чем больше различие между i и k. Обычно wki мал по сравнению со скоростью репродукции AiQi.
• 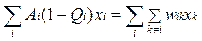 Если присутствуют все виды и все их возможные мутанты учитываются индексами i и k (i =1,N; k =1,N), то для ошибочных копий выполняется следующий закон сохранения:
Если присутствуют все виды и все их возможные мутанты учитываются индексами i и k (i =1,N; k =1,N), то для ошибочных копий выполняется следующий закон сохранения:
• 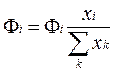 Добавление или удаление вида i любым способом, кроме химической реакции описывается членом Фi. Этот член задает индивидуальный поток или транспорт. С его помощью учитываются метаболические превращения. Суммарный поток Фt подсчитывается как нормированная сумма вкладов всех видов, поскольку в большинстве случаев каждый вид вносит вклад в суммарный поток в соответствии со своим изобилием:
Добавление или удаление вида i любым способом, кроме химической реакции описывается членом Фi. Этот член задает индивидуальный поток или транспорт. С его помощью учитываются метаболические превращения. Суммарный поток Фt подсчитывается как нормированная сумма вкладов всех видов, поскольку в большинстве случаев каждый вид вносит вклад в суммарный поток в соответствии со своим изобилием:
• 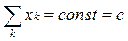 Суммарный поток в эволюционных экспериментах можно подобрать так, чтобы обеспечивалось поддержание воспроизводимых глобальных условий — например, таких, как постоянство суммарной плотности популяции:
Суммарный поток в эволюционных экспериментах можно подобрать так, чтобы обеспечивалось поддержание воспроизводимых глобальных условий — например, таких, как постоянство суммарной плотности популяции:
• 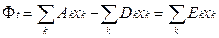 Для поддержания постоянной суммарной плотности популяции поток Фt необходимо постоянно изменять так, чтобы компенсировать избыточную общую продукцию, т. е.
Для поддержания постоянной суммарной плотности популяции поток Фt необходимо постоянно изменять так, чтобы компенсировать избыточную общую продукцию, т. е.
Величина Ei = Ai - Di является «избыточной продуктивностью» матрицы i. Благодаря уравнению сохранения, ошибки не влияют на вид этой суммы.
• 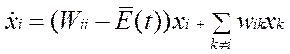 Можно ввести дополнительное ограничение - ограничение «постоянной общей организации»,- при котором как суммарное количество организованного материала, так и количество неорганизованного материала поддерживаются на постоянном уровне благодаря регуляции потоков. Обычно такое ограничение выполняется в эволюционных экспериментах, например в проточном реакторе. Тогда при условии соблюдения ограничения постоянной общей организации кинетические уравнения в сочетании с дополнительными условиями — принимают следующий вид:
Можно ввести дополнительное ограничение - ограничение «постоянной общей организации»,- при котором как суммарное количество организованного материала, так и количество неорганизованного материала поддерживаются на постоянном уровне благодаря регуляции потоков. Обычно такое ограничение выполняется в эволюционных экспериментах, например в проточном реакторе. Тогда при условии соблюдения ограничения постоянной общей организации кинетические уравнения в сочетании с дополнительными условиями — принимают следующий вид:
где величину
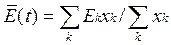 Wii = AiQi - Di
Wii = AiQi - Di
можно назвать внутренней селективной ценностью, а величину
— средней избыточной продуктивностью, которая является функцией времени. В момент, когда популяционные переменные xk(t) становятся постоянными (dxk(t) / dt =0), E(t) принимает стационарное значение, которое является метастабильным, т.к. зависит от полноты спектра мутантов, т.е. от того сколько мутантов присутствуют из всех возможных в системе. Для постоянных (т. е. не зависящих от времени) значений Wii и wik нелинейная система дифференциальных уравнений может быть решена.
КОНЦЕПЦИЯ КВАЗИВИДА
• В рассматриваемых формулировках, виды связаны друг с другом благодаря процессу мутирования, т.е. отдельный вид не является независимой единицей. Условие постоянства общей организации, а точнее - сохранение суммарной численности популяций всех матриц вынуждает все виды матриц («оригинальных» и мутантов) конкурировать друг с другом. Однако мутабильность допускает совместное существование (в некотором смысле кооперацию), особенно между близкородственными видами (т. е. видами i и k, для которых wik ≠0 и wki ≠0).
• 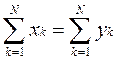 Реорганизуем систему следующим образом. Вместо разделения всей популяции на N видов матриц, введем новую систему N квазивидов. Каждому квазивиду соответствует популяционная переменная yi, которая является линейной комбинацией исходных популяционных переменных xi, причем их сумма, конечно, не изменяется:
Реорганизуем систему следующим образом. Вместо разделения всей популяции на N видов матриц, введем новую систему N квазивидов. Каждому квазивиду соответствует популяционная переменная yi, которая является линейной комбинацией исходных популяционных переменных xi, причем их сумма, конечно, не изменяется:
• 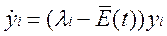 Каким образом выполнить это новое подразделение, подсказывает структура дифференциальных уравнений. Оно соответствует аффинному преобразованию системы координат, хорошо известному из теории линейных дифференциальных уравнений. Аффинное преобразование одного евклидова пространства в другое сохраняет параллельные прямые параллельными, а пересекающиеся – пересекающимися. Для трансформированных популяционных переменных yi получим новую систему уравнений:
Каким образом выполнить это новое подразделение, подсказывает структура дифференциальных уравнений. Оно соответствует аффинному преобразованию системы координат, хорошо известному из теории линейных дифференциальных уравнений. Аффинное преобразование одного евклидова пространства в другое сохраняет параллельные прямые параллельными, а пересекающиеся – пересекающимися. Для трансформированных популяционных переменных yi получим новую систему уравнений:
• 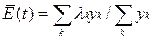 Применение этой процедуры к нелинейным уравнениям возможно благодаря тому, что член, ответственный за нелинейность, E(t), согласно уравнению, остается инвариантным при трансформации и теперь может быть выражен как среднее всех λi
Применение этой процедуры к нелинейным уравнениям возможно благодаря тому, что член, ответственный за нелинейность, E(t), согласно уравнению, остается инвариантным при трансформации и теперь может быть выражен как среднее всех λi
• Величины λi являются собственными значениями линейной динамической системы уравнений. Они, как и собственные векторы, которые связывают xi с уi, могут быть получены из матрицы, состоящей из коэффициентов Wii и wik.
• Физический смысл решений системы относительно переменных yi следующий: любой квазивид, собственное значение λi которого лежит ниже порога — среднего E(t) — будет вымирать, поскольку скорость изменения его численности отрицательна. И наоборот, численность каждого квазивида с λi выше порога будет расти. Однако, порог E(t) является функцией времени и будет расти.
• Рост будет происходить потому, что система благоприятствует квазивидам, которым соответствуют бОльшие собственные значения. Этот процесс будет продолжаться до тех пор, пока средняя продуктивность не сравняется с максимальным собственным значением и система не достигнет стационарного состояния. В этом стационарном состоянии в системе останется только один квазивид – тот, которому соответствует максимальное собственное значение, все остальные окажутся вымершими. Их популяционные переменные должны обратиться в ноль.
• Квазивид, как мы видим, понятие вполне теоретическое, полученное из математических моделей. Каков же физический смысл квазивида? Квазивид можно назвать «диким типом» популяции связанных друг с другом мутационным процессом молекул. В популяционной генетике обычно полагают, что дикий тип — это «стандартный» генотип, который в распределении генотипов в популяции соответствует оптимально адаптированному фенотипу. Однако лишь малая доля последовательностей полностью идентична той, которая считается диким типом.
• Большинство последовательностей являются копиями с одной или несколькими ошибками, и только среднее по распределению этих копий адекватно последовательности дикого типа. Численные эксперименты показывают, что стандартные копии (дикий тип) могут составлять меньше нескольких процентов всей популяции. Подавляющая часть популяции состоит из нестандартных копий (мутантов), более или менее далеких от дикого типа. При этом каждый конкретный мутант (нестандартная копия), по сравнению со стандартной копией, присутствует в этом распределении в очень малом количестве.
• Тогда полное распределение характеризуется некоей усредненной последовательностью, которая и определяет дикий тип. Квазивид, таким образом, представляет собой такое организованное распределение, которое характеризуется одной (или несколькими) усредненными последовательностями. Единственная усредненная последовательность имеется в единственном же случае - если стандартная копия явно доминирует. Явное доминирование выражается в том, что она имеет максимальную селективную ценность в пределах данного распределения.
• Те мутанты, у которых Wii очень близки к максимальным значениям, будут (в среднем) присутствовать в популяции в бОльших количествах. Эта неравномерность приведет к тому, что последовательность дикого типа будет «недоопределена» в некоторых позициях, т.е. некоторые символы в последовательности будут определяться неоднозначно. Два близкородственных мутанта могут иметь практически одинаковые селективные ценности, тогда оба они будут входить в квазивид с почти равными статистическими весами.
• Эти мутанты могут стать селекционно неразличимы, если значения Wii для них будут достаточно близки.
• Насколько близки друг к другу должны быть значения Wii для этих двух мутантов, чтобы они стали селекционно неразличимы, зависит от их «степени родства». Ясно, что соответствие Wii для отдаленно родственных последовательностей должно быть гораздо более точным, чем для копий с одной ошибкой.
КАТАСТРОФА МУТАЦИОННЫХ ОШИБОК
• Поскольку любой процесс репликации последовательности происходит с ошибками (мутациями), то это приводят к появлению квазивидов, представляющих из себя автономные группы последовательностей, мутационно переходящих друг в друга. Как было показано, устойчивость квазивидов - обязательное условие устойчивости ансамблей, в которые они входят. Кроме того, квазивиды, участвующие в циклических системах (гиперциклах, сайзерах) стабилизируются. Последовательность, составленная из наиболее вероятных символов в каждой позиции, называется "консенсусом" некоей группы последовательностей (квазивид в частности).
• Устойчивость консенсуса обеспечивает устойчивость квазивида.
•  Эйген и Шустер показали, что длина оптимальной матрицы (vm), средняя вероятность мутирования на позицию (1-) и параметр отбора матрицы (σт) связаны неравенством
Эйген и Шустер показали, что длина оптимальной матрицы (vm), средняя вероятность мутирования на позицию (1-) и параметр отбора матрицы (σт) связаны неравенством
• Будем считать матрицу, имеющую наибольший параметр отбора, оптимальной. В таком случае, возможные длины последовательностей оказываются ограниченными сверху величиной vmax. Нарушение этого неравенства называется "катастрофой мутационных ошибок". Можно выделить три режима эволюции по отношению к границе катастрофы ошибок.
• 1) Квазивид далек от границы катастрофы ошибок (1- << 1/vmax). В этом случае скорость эволюции низка, от интенсивности отбора зависит слабо. При стабилизирующем в квазивиде присутствуют несколько близких (родственных) последовательностей, в том числе и оптимальная, которая доминирует. При движущем отборе оптимальная последовательность достигается медленно.
• 2) Квазивид находится вблизи порога катастрофы ошибок (1- < 1/vmax). Тогда, в среднем, каждая копия в каждом поколении имеет одну ошибку. Однако порог не нарушен, и скорость эволюции близка к максимальной. Доля оптимальной последовательности мала. Консенсус становится "идеальной" характеристикой квазивида в том сиысле, что он существует и устойчив, но не обязательно совпадает с какой-либо реально существующей последовательностью.
• 3) Порог катастрофы ошибок нарушен
• (1- > lnσm/vmax). В этом случае у квазивида нет ни устойчивого консенсуса, ни стационарной структуры, он подвержен случайным флуктуациям. Фактически квазивид в этом случае просто не существует.
• Судя по всему, ограничения на длину последовательности в силу неточного воспроизведения, неоднократно имели место в ходе как добиологической, так и биологической молекулярной эволюции. Ясно, что на ранних этапах добиологической эволюции вероятность ошибок репликации должна была быть очень велика. Из экспериментальных данных известно, что ошибки комплементарного узнавания нуклеотидов без участия ферментов репликации, 1- ~10 -2.
• Соответственно, размер матриц ограничен vmax ~10 2, мономеров. При появлении ферментов репликации граница отодвигается до ~ 103 - 104. Возникновение репарационных механизмов коррекции ошибок репликации у прокариот увеличивает vmax до ~ 106 - 107 (E.coli и др.). Формирование у эукариот механизмов репарации повреждений генома, а также диплоидности, повышает эту границу до ~ 1012 -1014
• Такой подход имеет общеэволюционное значение - понятие консенсуса квазивида аналогично понятию нормы, постоянно используемому в популяционной и эволюционной генетике. Известны понятия видовой нормы, медицинской нормы, дикого типа и пр. Виды и популяции гетерогенны (если только это не лабораторные «чистые линии»), однако на основе совокупности наиболее часто встречаемых признаков видового уровня появляется представление о норме.
• Можно определить видовую норму как набор признаков, наиболее характерных для вида. Так называемая норма может быть идеальной конструкцией, т.е. реальных носителей нормы в популяции нет, но до тех пор пока она устойчива, мы можем говорить о существовании вида.
• Палеонтологи проводят оценки длительности существования видов основываясь на соображениях сохранение видовой нормы. Так, например, длительность существования видов животных и растений оценивается в
~ 1-30 млн лет. Далее появляются отличия от исходных форм по видовой норме морфологических признаков.
• Итак, динамика консенсуса квазивида является в некотором смысле молекулярной моделью динамики видовой и популяционной нормы. При этом норму можно рассматривать как консенсус изменяющихся признаков, которые характерны для рассматриваемого вида или популяции.
Теория нейтральной эволюции Кимуры.
Режимы макроэволюции.
• Детерминированные модели динамики популяций
• Стохастические модели динамики популяций
• Связь параметров микроэволюционного и макроэволюционного процессов
• Генетический груз и дилемма Холдейна
• Скорости макроэволюции и роль многоклеточности
ДЕТЕРМИНИРОВАННЫЕ МОДЕЛИ ДИНАМИКИ ПОПУЛЯЦИЙ
• Под динамикой популяции будем понимать изменение во времени частот аллелей в популяции. Рассмотрим менделевскую (наследование признаков происходит по законам Менделя), бесполую, диплоидную (организмы имеют двойной набор хромосом), панмиктическую (могут скрещиваться все со всеми) популяцию с одним диаллельным локусом (А1, А 2), где отбор осуществляется на уровне гамет.
• Пусть гаметы имеют приспособленности w1 и w2, соответственно, а разница приспособленностей s=w1-w2. Примем за p частоту аллеля А1. Тогда мы можем записать детерминированное уравнение динамики
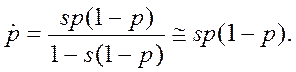 |
• У этого уравнения две тривиальные равновесные точки: = 0, = 1 и = 1, = 0.
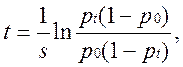 При s > 0 устойчива первая, в этом случае предпочтительный аллель А1, вытеснит аллель А2 из популяции, при s < 0 – устойчива вторая и в популяции победит аллель А2. Пусть р0 - начальная частота аллеля А1, а pt - частота в момент t, тогда прямое интегрирование уравнения дает
При s > 0 устойчива первая, в этом случае предпочтительный аллель А1, вытеснит аллель А2 из популяции, при s < 0 – устойчива вторая и в популяции победит аллель А2. Пусть р0 - начальная частота аллеля А1, а pt - частота в момент t, тогда прямое интегрирование уравнения дает
• Из чего видно, что время достижения частоты pt обратно пропорционально селективному преимуществу s адаптивного аллеля. Рассмотрим эту же задачу, но будем считать, что отбор осуществляется на уровне не гамет, а диплоидных особей. Обозначим приспособленности трех возможных генотипов A1A1, А1А2 и А2А2 через w11, w12 и w22 соответственно.
• 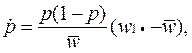 Соответствующее дифференциальное уравнение динамики для частоты аллеля А1 имеет вид
Соответствующее дифференциальное уравнение динамики для частоты аллеля А1 имеет вид
где
w1• = w11p + w12(1-p),
w2• = w21p + w22(1-p),
а
= w1•p+w2• (1-p) = w11p2 + 2w12p(1-p) + w22(1-p)2
Из полученных выражений видно, что, если w11 > w12 > w22, то устойчива равновесная точка = 1, = 0. В этом случае более приспособленный аллель А1 вытесняет аллель А2 из популяции. Если пересчитать приспособленность на гаметы, окажется, что аллель А1, в любой момент в среднем более адаптивен, чем А2:
w1• > w2•
• В случае w11 < w12 < w22 картина противоположная.
• 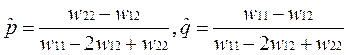 Если w11 < w12, и w22 < w12, то есть идет отбор в пользу гетерозигот (в генотипе присутствуют два разных аллеля), то будет устойчива нетривиальная равновесная точка, описывающая полиморфизм, с частотами аллелей
Если w11 < w12, и w22 < w12, то есть идет отбор в пользу гетерозигот (в генотипе присутствуют два разных аллеля), то будет устойчива нетривиальная равновесная точка, описывающая полиморфизм, с частотами аллелей
• При описании дизруптивного отбора, т.е. ситуации, когда приспособленность больше у гомозигот (в генотипе присутствуют два одинаковых аллеля), что выражается в неравенствах w11 > w12 и w22 > w12, полиморфная точка неустойчива и служит сепаратриссой – границей раздела двух зон устойчивости вокруг мономорфных точек
= 0, = 1 и = 1, = 0.
Описанные выше модели демонстрируют зависимость фазового портрета пространства частот от различных вариантов отбора особей с дифференциальной адаптивностью аллелей.
Теория нейтральной эволюции Кимуры.
Режимы макроэволюции.
• Детерминированные модели динамики популяций
• Стохастические модели динамики популяций
• Связь параметров микроэволюционного и макроэволюционного процессов
• Генетический груз и дилемма Холдейна
• Скорости макроэволюции и роль многоклеточности
• Под динамикой популяции будем понимать изменение во времени частот аллелей в популяции. Рассмотрим менделевскую (наследование признаков происходит по законам Менделя), бесполую, диплоидную (организмы имеют двойной набор хромосом), панмиктическую (могут скрещиваться все со всеми) популяцию с одним диаллельным локусом (А1, А 2), где отбор осуществляется на уровне гамет.
• Пусть гаметы имеют приспособленности w1 и w2, соответственно, а разница приспособленностей s=w1-w2. Примем за p частоту аллеля А1. Тогда мы можем записать детерминированное уравнение динамики
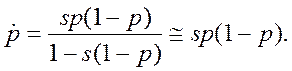 |
• У этого уравнения две тривиальные равновесные точки: = 0, = 1 и = 1, = 0.
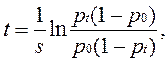 При s > 0 устойчива первая, в этом случае предпочтительный аллель А1, вытеснит аллель А2 из популяции, при s < 0 – устойчива вторая и в популяции победит аллель А2. Пусть р0 - начальная частота аллеля А1, а pt - частота в момент t, тогда прямое интегрирование уравнения дает
При s > 0 устойчива первая, в этом случае предпочтительный аллель А1, вытеснит аллель А2 из популяции, при s < 0 – устойчива вторая и в популяции победит аллель А2. Пусть р0 - начальная частота аллеля А1, а pt - частота в момент t, тогда прямое интегрирование уравнения дает
• Из чего видно, что время достижения частоты pt обратно пропорционально селективному преимуществу s адаптивного аллеля. Рассмотрим эту же задачу, но будем считать, что отбор осуществляется на уровне не гамет, а диплоидных особей. Обозначим приспособленности трех возможных генотипов A1A1, А1А2 и А2А2 через w11, w12 и w22 соответственно.
• Соответствующее дифференциальное уравнение динамики для частоты аллеля А1 имеет вид
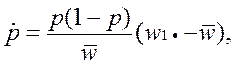 |
где
w1• = w11p + w12(1-p),
w2• = w21p + w22(1-p),
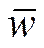 а
а
= w1•p+w2• (1-p) = w11p2 + 2w12p(1-p) + w22(1-p)2
Из полученных выражений видно, что, если w11 > w12 > w22, то устойчива равновесная точка = 1, = 0. В этом случае более приспособленный аллель А1 вытесняет аллель А2 из популяции. Если пересчитать приспособленность на гаметы, окажется, что аллель А1, в любой момент в среднем более адаптивен, чем А2:
w1• > w2•
• В случае w11 < w12 < w22 картина противоположная.
• 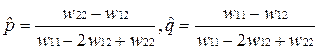 Если w11 < w12, и w22 < w12, то есть идет отбор в пользу гетерозигот (в генотипе присутствуют два разных аллеля), то будет устойчива нетривиальная равновесная точка, описывающая полиморфизм, с частотами аллелей
Если w11 < w12, и w22 < w12, то есть идет отбор в пользу гетерозигот (в генотипе присутствуют два разных аллеля), то будет устойчива нетривиальная равновесная точка, описывающая полиморфизм, с частотами аллелей
• При описании дизруптивного отбора, т.е. ситуации, когда приспособленность больше у гомозигот (в генотипе присутствуют два одинаковых аллеля), что выражается в неравенствах w11 > w12 и w22 > w12, полиморфная точка неустойчива и служит сепаратриссой – границей раздела двух зон устойчивости вокруг мономорфных точек
= 0, = 1 и = 1, = 0.
Описанные выше модели демонстрируют зависимость фазового портрета пространства частот от различных вариантов отбора особей с дифференциальной адаптивностью аллелей.
СТОХАСТИЧЕСКИЕ МОДЕЛИ ДИНАМИКИ ПОПУЛЯЦИЙ
• Те же задачи можно рассмотреть в стохастической постановке. Пусть имеется конечная менделевская панмиктическая популяция с постоянной эффективной численностью Ne = const. Изменение генетической структуры популяции происходит благодаря отбору гамет или особей.
• Единственный случайный фактор динамики популяции - биномиальный выбор 2Ne гамет при формировании следующего поколения. Данный популяционный фактор является одним из важнейших и может привести к серьезным последствиям. Стохастическим следствием случайного биномиального выбора гамет оказывается асимптотическая фиксация и утрата аллелей
• 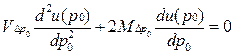 Если рассмотреть процесс стохастической динамики частот аллелей в популяции как аналог диффузионного процесса, то в этом приближении его можно описывать прямым и обратным уравнениями Колмогорова. В частности, при изучении вопроса об асимптотической фиксации аллелей обычно используют стационарное обратное уравнение Колмогорова
Если рассмотреть процесс стохастической динамики частот аллелей в популяции как аналог диффузионного процесса, то в этом приближении его можно описывать прямым и обратным уравнениями Колмогорова. В частности, при изучении вопроса об асимптотической фиксации аллелей обычно используют стационарное обратное уравнение Колмогорова
• В этом уравнении: и(р0) — условная вероятность асимптотической фиксации аллеля А1, если eго начальная частота равна р0 , — математическое ожидание изменения его частоты за поколение, a — дисперсия случайного изменения его частоты за поколение.
• Если предположить, что единственным случайным фактором в процессе жизни и размножения популяции является биномиальный выбор аллелей при формировании зигот следующего поколения, тогда
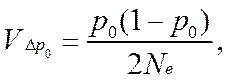 |
• а в качестве используются правые части соответствующих детерминированных уравнений динамики
• 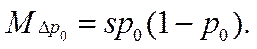 В случае гаметного отбора имеем
В случае гаметного отбора имеем
• 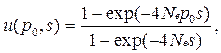 В таком случае, вероятность асимптотической фиксации аллеля А1 (решение уравнения Колмогорова) будет имеет вид
В таком случае, вероятность асимптотической фиксации аллеля А1 (решение уравнения Колмогорова) будет имеет вид
• Среднее время до фиксации t1(р0) выглядит достаточно сложно, поэтому приведем его только для некоторых существенных частных случаев.
• Например, рассмотрим ситуацию, когда аллель А1 нейтрален по отношению к А2 (т. е. s = 0), а начальная частота аллеля А1 р0 = 1/2 Ne, т. е. А1 – единственный в популяции (возник как мутация)
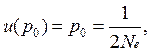 |
Тогда вероятность асимптотической фиксации аллеля А1
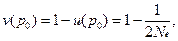 |
вероятность его случайной утраты
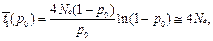 |
• среднее время до фиксации (исключая случаи утери аллеля)
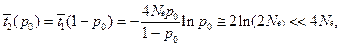 |
а среднее время до случайной утери
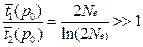 Соотношение этих величин
Соотношение этих величин
• Таким образом, для 2Ne ≈ 104, t1(р0) ≈ 2•104 поколений, t2(p0) ~ 18 поколений, а на одну фиксацию приходится ~2Ne ≈ 104 случайных утрат.
• Ясно, что вероятность фиксации адаптивного аллеля
u(p0,s) > u(p0),
а
(p0,s) < (p0) при s>0 и р0 = 1/2Ne
• В то время, как для инадаптивного аллеля соотношения противоположные:
u(p0,s) < u(p0),
(p0,s) > (p0) при s<0 и р0 = 1/2Ne
• 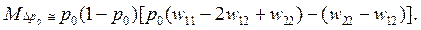 Рассмотрим стохастический вариант проанализированной выше детерминистической задачи об отборе на уровне генотипов особей. В этом случае
Рассмотрим стохастический вариант проанализированной выше детерминистической задачи об отборе на уровне генотипов особей. В этом случае
Величины же u(p0,s), t1(p0,s) можно подсчитать только численными методами.
• Диаллельный случай отбора генотипов может быть качественно сведен к анализу пересчитанных приспособленностей гамет. Поэтому задача о гаметном отборе, хотя она и крайне упрощена как в детерминистическом, так и в стохастическом вариантах, может служить ориентиром при проведении анализа задачи стохастической фиксации аллелей при эволюции макромолекул.
• В результате, можно сделать вывод, что микроэволюционные стохастические процессы в популяциях конечной численности относительно редко заканчиваются фиксацией одних аллелей и относительно часто потерей других. При этом стохастические флуктуации частот аллелей становятся существенными в течение количества поколений, сравнимых с эффективной численностью популяций.
t ≥ 2Ne
• Этот вывод – один из наиболее важных результатов теории нейтрального генетического дрейфа - теории нейтральной эволюции Кимуры. Его можно использовать (и используют) для объяснения механизмов эволюции макромолекул.
СВЯЗЬ ПАРАМЕТРОВ МИКРОЭВОЛЮЦИОННОГО И МАКРОЭВОЛЮЦИОННОГО ПРОЦЕССОВ
• Кимура считал, что завершенные процессы микроэволюции (фиксации замен и т. д.) можно считать элементарными событиями макроэволюции. Фактически ему первому удалось связать параметры микро- и макроэволюции, используя разработанную теорию нейтрального генетического дрейфа.
• Первичные структуры аллельных и гомологичных генов и белков различны, они имеют фиксированные замены отдельных мономеров. Эти фиксированные замены отдельных мономеров, которые не затрагивают остальных мономеров последовательности, являются элементарными макроэволюционными различиями. Однако же, аллели и гомологичные гены большей частью сохраняют сходство между собой.
• Аллели, фиксированные на промежуточных этапах макроэволюции, редко исчезает бесследно. Обычно аллели-потомки имеют в своей первичной структуре (последовательности) ранее фиксированные замены мономеров.
• Фиксация аллеля означает, что все аллели-потомки хранят в своей первичной структуре эти фиксированные замены. При этом в популяции могут существовать и дугие аллели – другие варианты гена.
• Определим макроэволюционный процесс как последовательность стохастических фиксаций замен, происходящих в предыдущих аллелях из-за мутаций. Большинство из возникающих замен теряется, поэтому полное число замен должно быть много больше количества фиксируемых. Тогда для i-го локуса скорость фиксации замен с адаптивным преимуществом s = w1 - w2 будет равна
• Ki(s) = 2Ne μi(s) u(p0,s)
• где в правой части стоит комбинация микроэволюционных параметров: Ne — эффективная численность популяции, μi(s) — плотность вероятности мутирования i -го гена в аллели с селекционным преимуществом s в единицу времени, а и(р0, s) — вероятность асимптотической фиксации уже возникшего аллеля,
• Эти микроэволюционные параметры определяют величину Ki(s), которая является важнейшим макроэволюционным параметром. Таким образом, скорость фиксации замен определяется мутационным процессом, отбором и интенсивностью генетического дрейфа.
Для одиночно возникшей нейтральной мутации
р0 = 1/2Ne, s = 0, и u(p0,s) = р0 = 1/2Ne,
откуда
Ki(0) = μi(0),
• т. е. скорость фиксации нейтральных мутаций определяется только скоростью их возникновения. Суммарная скорость фиксации всех адаптивных и нейтральных замен для i-го локуса будет
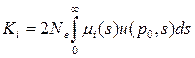 |
• Как показывают дополнительные исследования, дилемма Холдейна не возникает для нейтральных или почти нейтральных мутаций, т. е. отсутствуют ограничения на число одновременно эволюционирующих генов. В отличие от адаптивных мутаций выборочная случайная фиксация нейтральных аллелей не влияет на приспособленность и никак не изменяет скорость воспроизводства популяции.
Концепцию нейтральной эволюции Кимура и Ота сформулировали в виде 5 принципов:
- Скорость эволюции белка (число аминокислотных замен на позицию в год) остается почти постоянной пока трехмерная структура и функция серьезно не изменены.
- Более функционально значимые части молекулы белка (или целые молекулы) меняются путем замен медленнее, чем менее значимые.
- Замены меньше искажающие структуру и функцию макромолекул фиксируются в эволюции много чаще.
- Появлению гена с новой функцией обязательно должна предшествовать дупликация гена-предшественника.
- Дарвиновский отбор заведомо предпочтительных мутантов происходит в процессе эволюции редко, много чаще происходит случайная фиксация слабодефектных и нейтральных мутантов, а также удаление заведомо вредных.
ГЕНЕТИЧЕСКИЙ ГРУЗ И ДИЛЕММА ХОЛДЕЙНА
Введем меру интенсивности генетического отбора – величину, которая представляет собой отношение разности между средней приспособленностью популяции и приспособленностью оптимального генотипа к приспособленности последнего. Под оптимальным генотипом будем понимать наиболее приспособленный. Эта мера будет описывать генетический груз популяции. Чем ближе средний уровень приспособленности к оптимальному, тем меньше генетический груз.
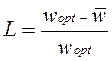 |
• Генетический груз L определяется следующим образом
где - средняя приспособленность популяции,
а - приспособленность оптимального генотипа.
• Собственно, под грузом можно понимать слабо приспособленные геномы, имеющие относительно большое количество «вредных» генов. Проблема существования и увеличения генетического груза оказалась одной из трудностей в развитии селективной теории генетики популяций и микроэволюции.
• Для очень большой диплоидной популяции со случайным скрещиванием, Холдейн показал, что при появлении полностью рецессивных вредных мутаций в аутосомном локусе, соответствующее снижение приспособленности равно скорости мутирования на гамету. Для полудоминантных мутаций груз равен скорости мутирования на зиготу.
• Причем груз почти не зависит от степени вредности мутаций. При других условиях, накладываемых на популяцию, локус и мутации, эти закономерности несколько модифицируются. Например, в случае очень малых популяций, в которых частоты мутаций дрейфуют от равновесных значений, оказывается, что в этих популяциях груз значительно выше, чем в больших
• Несколько упрощая формулировки, дилемма Холдейна заключается в том, что популяция с ограниченной скоростью воспроизводства не может быстро эволюционировать по многим генам одновременно.
• Холдейнпоказал математически, что даже в случае однозначного соответствия ген-признак в популяции не может заменяться одновременно свыше 12 генов «более приспособленными» аллелями без того, чтобы ее репродуктивная численность не стала исчезающее малой. Однако изучение природных популяций показало, что полиморфные популяции отличаются по очень большому числу аллелей, а вообще у организмов могут быть тысячи аллелей.
• Таким образом, согласно Холдейну, для поэтапной реализации таких различий природе потребовалось бы значительно больше времени, чем это в ряде случаев реально наблюдается. Дилемма Холдейна, демонстрирующая противоречие теории с экспериментальными данными, была частично разрешена лишь М. Кимурой, создавшего теорию нейтральной эволюции, которая основана на отказе от идеи исключительно селективной природы популяционных процессов.
СКОРОСТИ МАКРОЭВОЛЮЦИИ И РОЛЬ МНОГОКЛЕТОЧНОСТИ
Одноклеточные и многоклеточные организмы имеют разные характерные эффективные численности и среднее время жизни особей, т.е. характерную длительность поколения. Насколько многоклеточность особей влияет на скорость фиксации замен? Чтобы исследовать этот вопрос рассмотрим упрощенную модель
• Сравним две популяции:
• а) одноклеточную, имеющую эффективную численность 2 Ne, длительность поколения τ, темп мутирования в класс адаптивности (s) на поколение μτ(s), вероятность асимптотической фиксации и(р0, s) и скорость фиксации замен за поколение K τ (s).
• Будем исходить из положения, что все фиксируемые мутации принадлежат какому-то одному классу адаптивности (s). Тогда
Kτ(s) = 2Ne μτ(s) u(p0,s)
• б) многоклеточную, имеющую соответствующие параметры
2 Ne*, τ*, μ*τ* (s*), и*(р*0, s*),
для нее
K*τ*(s*) = 2N*e μ*τ*(s*) u*(p*0, s*).
• Будем считать, что длительность поколения τ* = т τ, где т — число клеточных поколений по генеративной линии в ходе онтогенеза. Также предположим для простоты, что мутационные параметры клеток и одноклеточных, и многоклеточных организмов за цикл репликации одинаковы.
• Поскольку многоклеточные особи имеют время развития в m раз больше одноклеточных, то темп их мутирования в класс адаптивности
(s = s*) за поколение μ*τ*(s)= m μτ (s). Поделив соответствующие выражение друг на друга, получаем
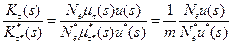 |
• Скорости фиксации замен выражаются через время, измеряемое в поколениях. Длительность поколения для одноклеточных и многоклеточных различна: τ и τ* соответственно. Однако мы можем пересчитать скорости фиксации замен на одинаковые единицы времени, например на время τ.
• 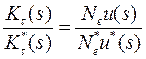 Для этого достаточно взять K *τ (s) = K* τ * (s)/m. В результате имеем
Для этого достаточно взять K *τ (s) = K* τ * (s)/m. В результате имеем
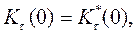 Предположим, что фиксирующиеся мутации нейтральны в обеих популяциях, т.е. s = s* = 0. Тогда и = pо = 1/2Ne,
Предположим, что фиксирующиеся мутации нейтральны в обеих популяциях, т.е. s = s* = 0. Тогда и = pо = 1/2Ne,
и* = pо * = 1/2N*e
т. е. скорости фиксации нейтральных замен в популяциях одноклеточных и многоклеточных особей одинаковы.
 Предположим, что фиксирующиеся мутации одинаково адаптивны: и ≈ 2s = u* ≈ 2s*.
Предположим, что фиксирующиеся мутации одинаково адаптивны: и ≈ 2s = u* ≈ 2s*.
В этом случае и не зависит от Ne, поэтому получаем
По данным популяционной экологии, для популяций высших млекопитающих принимается Ne≈ 104, тогда как для прокариот и одноклеточных Ne > 108
Т. е. скорость фиксации адаптивных замен в единицу времени у многоклеточных на много порядков ниже, чем у одноклеточных.
Даже, если допустить некоторые различия между параметрами мутирования на репликацию, а также между и и и*, то этот вывод сохранится.
• Полученный результат достаточно прост, однако он демонстрирует чрезвычайно важное стохастическое свойство развития живых форм на Земле. А именно, при переходе от одноклеточных к многоклеточным происходит резкое замедление темпа фиксаций именно адаптивных мутаций при сохранении прежнего темпа фиксаций нейтральных замен. Это замедление связано с резким падением численности популяций, замедлением воспроизведения и т. п.
• Видимо, для компенсации этого существенного ограничения (в том числе) многоклеточные системы приобрели новые или усилили старые (по сравнению с одноклеточными) механизмы изменчивости и комбинаторики генов, мРНК и белков. К этим механизмам можно отнести дублирование генов, интрон-экзонную структуру, транспозоны, множественные повторы, альтернативный сплайсинг, множественные старты трансляции, микроРНК, модификацию белков и т.д.
• Ясно, что следствия резкого замедление темпа фиксаций именно адаптивных мутаций в многоклеточных организмах по сравнению с одноклеточными должны особенно сильно проявляться при их непосредственном контакте - например в экосистемах «паразит — хозяин».
МОЛЕКУЛЯРНАЯ КОЭВОЛЮЦИЯ В СИСТЕМЕ «ФАГ - БАКТЕРИЯ»
• Существование огромного потенциального разнообразия эквифункциональных вариантов генов и белков определяет возможность одного из режимов эволюции — синонимического дрейфа, т. е. случайной фиксации изменений структуры макромолекул без изменения их функции.
• Учет неизбежной смены условий существования популяций (смены экологических ниш) и связанных с этим изменений критериев отбора привел к представлению, что адаптивность (приспособленность) аллелей в конкретных нишах не обязательно коррелирует с глобальной (длительной) адаптивностью.
• В результате возможны следующие варианты макроэволюционного процесса: а) локально адаптивный, но глобально не направленный (когда адаптивность предыдущего аллеля в соответствующей экологической нише не соответствует другой нише); б) локально адаптивный и глобально направленный процесс (когда при смене аллелей и переходе от ниши к нише функция гена в целом совершенствуется, приобретаются новые, глобально адаптивные свойства).
• Основная трудность в проверке и использовании формулы Кимуры применительно к природным популяциям состоит в том, что неизвестно распределение по классам приспособленности, а также, что имеются лишь очень смутные представления о последовательности смены экологических ниш, которая влияет на микроэволюционные параметры: эффективная численность популяции, классы приспособленности и т.д.
• Однако для экосистемы фаг — бактерия такие оценки, хотя и приблизительно, можно сделать. Коэволюционирующая экосистема «вирулентный фаг - бактерия» отражает реальную эволюционную ситуацию, когда экологические условия, принципы отбора и способы размножения популяций фага и бактерии предельно просты и измеримы и эволюционный процесс протекает существенно быстрее, но принципиальные особенности процесса макроэволюции сохранены.
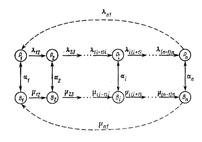
• Схема модели коэволюции контактирующих белков в системе фаг-бактерия. Обозначения: В1, В2,…, Вi, …, Вn – ряд штаммов бактерий, возникающих один из другого за счет мутирования в локусе белка рецепции; Р1, Р2,…, Рi, …, Рn - соответствующий ряд штаммов фага, вирулентных к бактериям с тем же номером. Пара <Bi, Pi > (i = 1,n) – обычная система «хищник-жертва»; mi(i+1) – скорость мутирования из i-го штамма бактерии в (i+1)-й штамм; l i(i+1) – соответствующая скорость мутирования у фагов; ai – константа скорости адсорбции i-го фага на i-й бактерии.
• В центре экологических отношений двух популяций (фага и бактерии) оказывается их способность к взаимной рецепции — адсорбции, которая реализуется путем специфического соответствия двух белков: белка рецепции у бактерий и белка адсорбции у фагов. Элементарными событиями макроэволюции естественно считать возникновения и фиксации замен нуклеотидов и аминокислот, поскольку это есть фиксация аллелей соответствующих генов в популяциях клеток E.Coli и бактериофага. Замена аминокислоты в белке рецепции нарушает связывание фага с бактерией, адекватная замена аминокислоты в белке адсорбции восстанавливает паразитические возможности фага.
МОЛЕКУЛЯРНАЯ КОЭВОЛЮЦИЯ В СИСТЕМЕ «ФАГ - БАКТЕРИЯ»
• Как показывает исследование соответствующих математических моделей, эволюция данной экосистемы складывается из колебаний концентраций бактерии и фага (с очень большой амплитудой), при этом в каждом следующем цикле участвуют новые мутантные штаммы жертвы-бактерии и хищника-фага.
•
Кинетика концентраций фага и бактерии в детерминированной модели
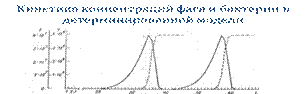
• Эволюционная смена пар мутантных форм в экосистеме фаг-бактерия. x0=1, y0=1; α =0.3•10-7, β =0.3, частоты мутантов: 10-5 для бактерий, 10-7 для фагов. По оси абсцисс отложено время (t), измеряемое в числе поколений, звездочками отмечены моменты появления мутантов.
• Период колебаний на графике составляет 21—22 поколения — это и есть среднее время фиксации мутаций. С учетом сказанного выше его можно поделить на два периода: 1) период «ожидания» (17—18 генераций), в течение которого мутантные клетки, имея низкую частоту, «ожидают» того момента, когда экологическая ниша изменится таким образом, что они окажутся предпочтительнее предшествующего варианта, и 2) период собственно ф и к с а ц и и, длящийся всего три-четыре генерации, когда за счет тотального лизиса исходного штамма мутантные клетки автоматически повышают свою частоту до 1.
• Это и есть модель локально адаптивной, но глобально ненаправленной коэволюции. При этом скорость процессов настолько велика (на фиксацию нового аллеля требуется ~25 поколений, т.е. около суток), что по фактическому масштабу времени это типичный микроэволюционный процесс, хотя по сути, как уже говорилось, - макроэволюционный.
• При дефиците ресурса, потребляемого бактерией для своего роста, модель «вирулентный фаг - бактерия» становится коэволюционно неустойчивой. Однако систему может "спасти" от вырождения приобретение фагом определенного адаптивного усложнения своей молекулярно - генетической системы управления - приобретения свойства умеренности. «Умеренность» означает появление у фага механизма выбора между литическим (убивающим бактерию) и лизогенным (встраиванием генома фага в хромосому бактерии) режимами развития, который как раз и связан с оценкой уровня ресурсов размножения бактерии.
• Исследование условий существования и устойчивости состояний равновесия системы «умеренный фаг - бактерия» позволяет сделать следующие выводы.
• При недостаточном обеспечении бактерий ресурсом реализуется устойчивое лизогенное состояние равновесия, когда в зависимости от начальных концентраций фагов и бактерий имеются либо только лизогенные клетки, либо смешанная популяция из здоровых и лизогенных клеток. В обоих случаях благодаря интеграции генома фага с геномом бактерии сохраняется целостность системы как таковой.
• Напротив, если система хорошо снабжается ресурсом, можно гарантировать существование и устойчивость лишь одного равновесного состояния – литического, при котором фаг убивает (лизирует) всю популяцию бактерий. Литическое состояние не означает, однако, гибель системы, поскольку именно при богатом ресурсе есть возможность коэволюции за счет возникновения новых мутантных пар фаг-бактерия
• Нетривиальное равновесие, когда в системе имеются и зараженные фагом, и здоровые бактерии, и свободные фаговые частицы, существует в среднем интервале значений ресурса. Очевидно, что если этот интервал узок, нетривиальное состояние равновесия стуктурно неустойчиво.
• Таким образом, в основе развития системы "умеренный фаг- бактерия" на популяционном уровне лежит триггерный механизм - система имеет два устойчивых состояния: лизогенное при дефиците ресурса и литическое при его избытке. В лизогенном состоянии система не вырождается благодаря сохранению паразита в виде профага, а в литическом - за счет гарантированного возникновения новой пары мутантных штаммов-антагонистов, т.е. коэволюции.
• Невырождения данной системы можно добиться и «более простым» способом: введя проток всех компонент через резервуар. Исследование системы, описывающей динамику одной пары штаммов вируса и бактерии, показывает, что при биологически реалистичных значениях параметров системы наиболее вероятным устойчивым состоянием является нетривиальное равновесие с сохранением и фага, и бактерии, т. е. с невырождением системы как таковой.
• Изучение процесса появления мутантов по белкам рецепции – адсорбции показывает, что наличие протока приводит к полиморфизму как наиболее вероятному состоянию системы фаг — бактерия. В этой связи отметим, что фактически проточная модель фаг — бактерия представляет собой интересный пример зависимости вероятности фиксации мутаций в одних генах (определяющих эффективность адсорбции) от мутаций в других генах (определяющих скорость воспроизведения).
• Реальные системы могут, очевидно, быть попеременно то проточными, то непроточными. Образно говоря, ситуация напоминает смену сухого сезона сезоном дождей. «Сухой сезон» отвечает изолированному резервуару, «сезон дождей» — проточному. Им соответствует качественное изменение режимов эволюции. В изолированном резервуаре при избытке ресурсов возможна устойчивая коэволюция популяций вирулентного и умеренного фага и бактерии.
• При ограниченном ресурсе — лизогенное состояние системы умеренный фаг — бактерия. В проточном резервуаре коэволюция фага и бактерии невозможна — устанавливается сбалансированный полиморфизм пар мутантных вариантов. А в целом такая смена внешних условий переводит систему из микроэволюционно стабильного состояния полиморфизма в стабильный режим коэволюции и обратно
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 855; Нарушение авторских прав?; Мы поможем в написании вашей работы!