КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Синтез і розпад жирів
|
|
|
|
Обмін ліпідів
Метаболічні шляхи, пов'язані з синтезом і розпадом тригліцеридів, у тварин і рослин багато в чому подібні, але є і відмінності.
Синтез насичених жирних кислот з парним числом вуглецевих атомів відбувається шляхом послідовного додавання двовуглецевих залишків до ацильного ланцюга, що бере свій початок від ацетил-СоА. Донором двовуглецевих одиниць є малоніл-СоА.
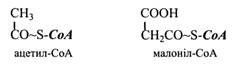
Цей процес каталізується поліферментним комплексом, який називають синтетазою жирних кислот. Кінцевим продуктом її дії є С16-пальмітинова кислота.
У рослинах є ще дві синтетази жирних кислот – складні ферментні системи. Одна з них подовжує вуглецевий ланцюг кислоти від С16 до С18, друга – від С18 до С30 і більше.
Синтетази жирних кислот у рослинних клітинах локалізовані в цитозолі, у стромі хлоропластів та в сферосомах.
Перетворення насичених жирних кислот у ненасичені відбувається за допомогою десатуразної ферментної системи, що вводить подвійний зв'язок у молекулу насиченої кислоти незалежно від довжини її вуглецевого ланцюга. Однак найчастіше субстратом десатурази є С18-стеаринова кислота, а продуктом – олеїнова. У реакції використовується NАDРН і O2.
Десатурази рослин мають ряд відмінностей від тваринних. Вони локалізовані в матриксі цитоплазми та в сферосомах, а тваринні – у мембранах ЕР. Рослинні десатурази можуть використовувати для реакцій електрони безпосередньо від фотосистеми при фотосинтезі.
Подальше перетворення мононенасичених жирних кислот у ди- і триненасичені ще недостатньо вивчено. Однак припускають участь у цьому процесі спеціальних десатураз. Відомо також, що в цих реакціях потрібні NАDРН і O2.
Десатурази у тварин утворюють поліненасичені жирні кислоти, вводячи подвійні зв'язки між уже існуючим подвійним зв'язком і карбоксильною групою, а у рослин – між наявним і кінцевою метильною групою.
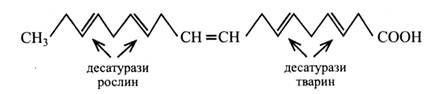
Завдяки цьому в рослин олеїнова кислота поступово перетворюється у лінолеву та ліноленову, а тварини ці кислоти синтезувати не можуть і повинні одержувати їх з їжею. Дія ж десатурази на ліноленову кислоту у тварин веде до утворення арахідонової кислоти, що має 4 подвійних зв'язки.
Недостатню вивченість механізму десатурації олеїнової кислоти в рослинах пояснюють тим, що ферменти пов'язані з мембранами ЕР, хлоропластів та сферосом і тому важко піддаються виділенню й очищенню.
Жирні кислоти, що відщеплються при гідролізі тригліцеридів під дією ліпази, далі підлягають β-окисленню з утворенням двовуглецевих фрагментів – ацетил-СоА. Цей процес подібний у рослин і тварин. Ферменти, що каталізують β-окислення жирних кислот, зосереджені в мітохондріях рослин.
Ацетил-СоА, що утворився при β-окисленні може піддаватися далі різним перетворенням. Основний шлях – це повне окислення до СO2 і Н2O у циклі Кребса. При цьому виділяється велика кількість енергії (12 молекул АТР). Частина молекул ацетил-СоА може знову використовуватися у синтезі жирних кислот або у синтезі інших сполук – каротиноїдів, стероїдів, ефірних олій і смол. Крім того, ацетил-СоА може йти на утворення вуглеводів, включаючись у ланцюг реакцій, які називають гліоксилатним циклом. Процес перетворення жирів у вуглеводи зветься глюконеогенезом.
Як зазначалося, багато рослин як запасні речовини у насінні відкладають жири (олії). Оскільки вони не розчиняються у воді, то і не можуть транспортуватися по рослині. Тому при проростанні насіння олійних рослин із жирів утворюються цукри, які й використовуються у живленні проростків. Це відбувається у процесі глюконеогенезу, через гліоксилатний цикл, що протікає у мікротільцях – гліоксисомах. Вони утворюються та функціонують у насінні олійних рослин при проростанні. Коли проросток зеленіє та переходить до автотрофного живлення, гліоксисоми поступово зникають.
У гліоксисоми надходять вільні жирні кислоти. Тут при дії комплексу всіх необхідних ферментів вони піддаються β-окисленню, яке дає велику кількість ацетил-СоА, що вступає в гліоксилатний цикл.
Він складається із шести реакцій. Чотири з них подібні до реакцій циклу трикарбонових кислот, а дві – специфічні тільки для цього циклу і в інших процесах обміну не зустрічаються.
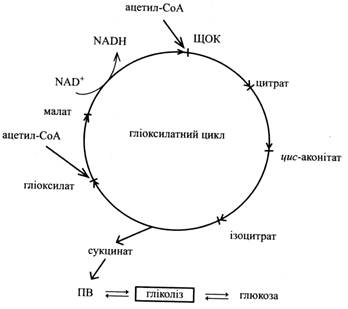
Схема гліоксилатного циклу и глюконеогенезу
Перші три реакції гліоксилатного циклу ідентичні таким же реакціям циклу трикарбонових кислот. Спочатку ацетил-СоА конденсується з оксалоцетатом з утворенням цитрату під дією цитратсинтази. Цитрат потім ізомеризується в ізоцитрат під впливом аконітат-гідратази. Проміжною сполукою в цій реакції є цис-аконітат.
Четверта реакція характерна тільки для гліоксилатного циклу: під дією ферменту ізоцитрат-ліази молекула ізоцитрату розщеплюється на сукцинат і гліоксилат. Сукцинат виходить з циклу і далі перетворюється у вуглеводи, а гліоксилат вступає в наступну реакцію циклу.
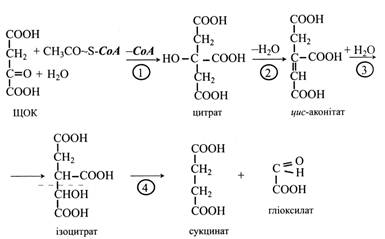
Гліоксилат реагує з ще однією молекулою ацетил-СоА, утворюючи малат. Малат-синтаза, що каталізує цю реакцію, так само специфічна тільки для гліоксилатного циклу, як і ізоцитрат-ліаза. В останній реакції циклу малат під дією малат-дегідрогенази за участю NАD+ окислюється до щавелеоцтової кислоти (ЩОК). Ця реакція характерна і для останнього етапу ЦТК.
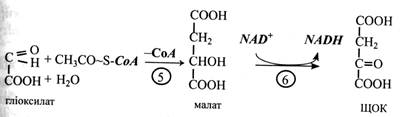
Продукт гліоксилатного циклу – сукцинат залишає гліоксисому, тому що в ній немає ферментів для його подальшого перетворення, і переходить у мітохондрію. Тут під впливом ферментів ЦТК він перетворюється на ЩОК. Спочатку, за дії сукцинат-дегідрогенази, сукцинат перетворюється на фумарат. Останній приєднує воду під впливом фумарат-гідратази, і утворюється малат. Малат-дегідрогеназа окислює малат до оксало-ацетату. При декарбоксилюванні останнього за участю АТР виникає ФЕП.

ФЕП входить у гліколіз і в процесі його обертання перетворюється на глюкозу.

Такий шлях глюконеогенезу в рослинах: від β-окислення жирних кислот, через гліоксилатний цикл, перетворення сукцинату та обертання гліколізу.
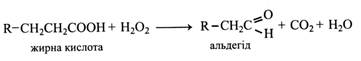
У той час як у тварин розпад насичених жирних кислот відбувається виключно шляхом β-окислення, рослини мають ще один шлях розщеплення жирних кислот – α-окислення. При цьому відбувається окислення α-вуглецевого атому жирної кислоти, що призводить до видалення карбоксилу у вигляді СO2 й утворення кислоти, вуглецевий ланцюжок якої коротше вихідної на один атом.
Кожне обертання α-окислення включає дві реакції. Перша рeакція відбувається під впливом пероксидази жирних кислот і за участю перекису – Н2O2. Продуктами реакції є альдегід, що містить на один вуглецевий атом менше, ніж кислота, вуглекислий газ і вода. Звичайно α-окислення відбувається в мікротільцях – пероксисомах, де в рослинній клітині утворюється перекис, необхідний у цьому процесі. Перекис звичайно виникає при окисленні гліколату під дією гліколат-оксидази.
У другій реакції відбувається окислення альдегіду. Тут діє фермент альдегід-дегідрогеназа за участю NaD+ і води.
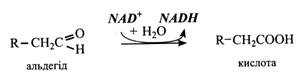
Фізіологічна роль α-окислення у рослині ще цілком не з'ясована. Припускають, що в результаті цього процесу утворюються жирні кислоти з непарним числом вуглецевих атомів. Крім того, обговорюється можливість окислення таким шляхом рідкісних жирних кислот, у яких біля β-вуглецевого атома є який-небудь замісник (-СН3, -ОН), що виключає можливість розщеплення даної кислоти звичайним шляхом β-окислення. Після α-окислення вуглець із замісником переміщується у α-положення і не заважає дії ферментів β-окислення.
Енергетично α-окислення майже в 3 рази менш ефективне, ніж β-окислення: у першому випадку утворюється лише 6 АТР проти 16 у другому. Хоча при α-окисленні відбувається менше реакцій і не потрібна активація жирної кислоти за допомогою Co А.
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 7041; Нарушение авторских прав?; Мы поможем в написании вашей работы!