КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Сукцессия сибирского темнохвойного леса после лесного пожара
|
|
|
|
| Время, лет | 15-25 | 25-50 | 75-100 | 100-150 | 125-175 | 150-200 | 200-250 |
| Фаза сукцес-сии | Луг | Кус-тар-ник | Березо-осино-вый лес | Смешан-ный лес | Сосно-вый лес | Сосново-кедро-вый лес | Кедрово-пихто-вый лес |
Конечным результатом сукцессии являются более медленно развивающиеся экосистемы, котоpые получили название климаксовых. Климаксовые экосистемы - относительно устойчивые фазы сукцессии, наиболее соответствующие экологическим характеристикам данной местности в определенный период геологического времени. Примерами таких систем в жарком и сухом климате являются пустыни, в жарком и влажном - тропические леса.
Как правило, сукцессия характеризуется прогрессивными процессами: формируется почва, развивается растительный покров, возрастает продуктивность экосистемы. Смена биоценозов сопровождается увеличением их видового разнообразия.
Сукцессионная смена биогеоценозов происходит в строгой последовательности, нарушение которой хозяйственным вмешательством, не всегда ведет к желаемому результату. Так, вырастить хвойный лес, который дает сырье для целлюлозно-бумажной промышленности, минуя фазу лиственного леса, для лесохозяйственников представляет немалые трудности. Таким обpазом, эксплуатируя природные ресурсы, человек должен знать и учитывать закономерности развития естественных экосистем.
2.3. Устойчивость экосистем
Устойчивость экосистемы - способность экосистемы и ее отдельных частей противостоять колебаниям внешних факторов и сохранять свою структуру и функциональные особенности. Например, в экосистеме количество осадков понижается на 50 % по сравнению со среднегодовыми значениями, но продукция растений уменьшается при этом только на 25 %, а численность популяции растительноядных организмов - всего на 10 %. Относительное затухание колебаний в среде по мере их прохождения по пищевым цепям служит мерой внутренней устойчивости экосистемы. Устойчивость экосистем обусловлена сбалансированностью потоков энергии и круговоротами веществ, а также наличием в экосистемах развитых информационных сетей, включающих потоки физических и химических сигналов, связывающих все части системы и управляющих ею как одним целым.
Можно считать, что экосистемы имеют кибернетическую природу, хотя в отличие от созданных человеком управляемых систем, ее управляющие функции сосредоточены внутри. Степень стабильности зависит от жесткости окружающей среды и от эффективности внутренних управляющих механизмов.
Выделяют два типа стабильности экосистем: резистентная устойчивость и упругая устойчивость. Резистентная устойчивость - это способность оставаться в устойчивом состоянии под нагрузкой, а упругая устойчивость - способность быстро восстанавливаться. Эти два типа стабильности связывает обратная зависимость.
Сравним элементарные схемы управления с особым, как в механическом устройстве, внешним управлением (рис. 6, а) и экосистему с регуляцией входящих в нее подсистем (рис. 6, б).
В обоих случаях управление основано на обратной связи, которая осуществляется, когда часть сигналов с выхода поступает на вход. Если обратная связь положительна, то значение управляемой переменной возрастает. Для того, чтобы осуществлять контроль, например, избегать перегрева в помещении (по первой схеме) или перенаселения (по второй схеме), необходима и отрицательная обратная связь, уменьшающая величину на входе. Для контроля перегрева в помещении необходимо задать некую постоянную температуру, с учетом которой термостат будет управлять калорифером. В экосистемах в результате взаимодействия круговоротов веществ и потоков энергии, а также сигналов положительной и отрицательной обратной связи от подсистем, возникает саморегулирующийся гомеостаз или состояние равновесия, без регуляции извне. Выявить кибернетическую природу экосистемы трудно, так как компоненты экосистемы связаны в информационные сети различными физическими и химическими агентами подобно тому, как нервная или гормональная системы связывают в одно целое части организма.
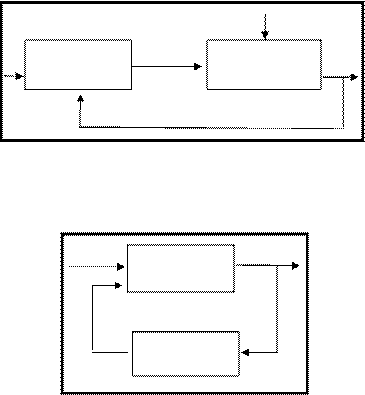 |
Заданные
величины


Управляющий Управляемая Выход
элемент субсистема
а)
Вход Первичная Выход
субсистема
Вторичная
субсистема
б)
Системы с обратной связью
Кроме систем обратных связей стабильность в экосистемах обеспечивается избыточностью функциональных компонентов. Например, если в экосистеме имеется несколько видов зеленых растений, каждое из которых характеризуется своим диапазоном температур, то скорость фотосинтеза в целом при колебаниях температур, может оставаться неизменной.
Гомеостатические механизмы функционируют в определенных пределах, за которыми уже ничем не ограничиваемые положительные обратные связи приводят системы к гибели. По мере нарастания стресса система, продолжая оставаться управляемой, может оказаться неспособной к возвращению на прежний уровень и занять другое состояние равновесия. Но в этом случае даже небольшие последующие нарушения могут иметь далеко идущие последствия. Надежный гомеостатический контроль устанавливается только после длительного, эволюционного периода времени. Например, природные экосистемы по сравнению с агроэкосистемами имеют более совершенный контролирующий механизм, поэтому колебания в них менее резки.
Степень стабильности, достигаемая конкретной экосистемой, зависит не только от ее развития, эффективности внутренних управляющих механизмов, но и характера окружающей среды и сложности самой экосистемы. Как правило, в благоприятной физической среде экосистемы имеют тенденцию становиться сложнее, а функциональная сложность увеличивает стабильность системы.
Рассмотрим два вида стабильности экосистем, резистентную устойчивость - способность экосистемы сопротивляться нарушениям, поддерживать свою структуру и функцию, и упругую устойчивость - способность восстанавливаться после того, как структура и функция были нарушены (рис. 7). Как правило, оба типа устойчивости экосистемам не удается развить в равной степени. То есть, если система обладает одним видом устойчивости, то практически не обладает другим. В настоящее время человек способен изменять функционирование экосистем, но в основном в своей деятельности человек наращивал эксплуатацию природных ресурсов, что уже привело к снижению качества человеческой жизни и нарушениям в окружающей среде.
 |
Возмущение
Диапазон нормального
функционирования
Мера Область
устойчивости устойчивости
Мера упругости
Время
Резистентная и упругая устойчивость экосистем
Поскольку глобальной экосистемой является биосфера Земли, остановимся на пределах ее устойчивости. Как отмечает К.Я. Кондратьев, в биосфере при отсутствии антропогенных возмущений потоки вещества за счет синтеза и разложения органических веществ естественной биотой почти совпадают, и это делает окружающую среду устойчивой в геологических масштабах времени. Иными словами, биосфера подчиняется принципу Ле-Шателье: при возникновении внешних возмущений, нарушающих состояние окружающей среды, в биоте должны возникать процессы, компенсирующие это возмущение. Однако воздействие человека на природные экосистемы привело к тому, что они начали терять способность к компенсации этих возмущений. Подсчитано, что биосфера может компенсировать возмущения, производимые человечеством, если доля ее продукции, потребляемая человеком, не превышает 1 %. Результаты новейших исследований показали, что действие принципа Ле-Шателье в биоте континентов оказалось нарушенным, как только доля потребления человеком продукции биосферы превысила порог допустимого воздействия (около 10 %).
Социально-экономическое развитие общества пришло в явное противоречие с ограниченными ресурсовоспроизводящими и жизнеобеспечивающими возможностями биосферы, происходит истощение естественных ресурсов суши и океана, безвозвратная потеря различных видов растений и животных, загрязнение окружающей среды, упрощение и деградация экосистем.
2.4. Популяция, ее структура и динамика
Наряду с определением экологии как науки об экосистемах на меньшее право на существование имеет определение экологии как науки о популяциях. Популяционный подход концентрирует свое внимание на отдельных видах. Чаще всего - это виды, имеющие важное хозяйственное значение: вредители сельского хозяйства, объекты промысла, переносчики болезней и т.д., но иногда это и просто массовые виды или виды редкие, нуждающиеся в охране. Популяция - это совокупность всех особей одного вида, входящих в конкретную экосистему и соответственно населяющих занимаемую этой экосистемой территорию или акваторию.
При описании популяции и ее количественном изучении используются обычно две группы количественных показателей: статические и динамические. Статические показатели характеризуют состояние популяции в какой-то определенный момент времени, динамические характеризуют процессы, протекающие в популяции за некоторый промежуток времени.
К статическим показателям популяции относят общую численность, плотность, различные показатели популяционной структуры (возрастной, размерной, половой).
В ходе некоторых экологических, а особенно эколого-генетических исследований возникает вопрос о том, какова общая численность особей в популяции. Оценка общей численности особенно важна для исчезающих, занесенных в Красную книгу видов животных или растений. Как правило, численность нетрудно определить, если имеешь дело с лабораторной популяцией достаточно крупных объектов (например, мучных жуков в банке).
Оценить общую численность природных популяций гораздо сложнее. Как правило, для оценки численности используют фотографический метод и метод мечения. Например, фотографическим методом можно подсчитать птиц в колониях, а методом мечения - рыб во внутренних водоемах. В подавляющем большинстве случаев эколог не в состоянии определить непосредственно общую численность популяции, а вынужден ограничиваться отбором отдельных проб и подсчетом особей в пробах. Величина, которую он при этом определяет, строго говоря, уже не является численностью, но является численностью, приходящейся на единицу пространства, то есть плотностью.
Для каждой группы организмов существуют свои способы оценки и выражения плотности. Так, например, при изучении травянистых растений используют квадратные или круглые рамки, которые кладут на землю, а затем подсчитывают все попавшие внутрь рамки растения. Иногда оценивают не собственно плотность растений, а покрытие площади в процентах.
Плотность популяции наземных растений и животных выражают обычно на единицу площади поверхности, а плотность популяций планктонных организмов - на единицу объема водной среды или же на единицу площади поверхности воды.
Из всего многообразия пространственного распределения популяции можно выделить три основных: случайное, регулярное и пятнистое. На практике для различения типов пространственного распределения пользуются различными статистическими методами. Самый простой из них следующий - это оценить дисперсию наблюдаемого распределения. Поясним это на примере. Пусть имеется популяция какого-либо вида травянистых растений, занимающих большой луг. Для оценки средней плотности помещают стандартную рамку определенной площади, а затем подсчитывают растения интересующего вида. Получив данные по некоторому количеству таких рамок, считают среднее число особей, приходящихся на одну площадку, М, (это и будет оценка средней плотности), а также определить дисперсию s2, которая высчитывается как средний квадрат каждого отклонения от значения средней плотности. При истинно случайном распределении дисперсия равна среднему, т.е. М = s2. При регулярном распределении дисперсия меньше среднего, а при пятнистом распределении - дисперсия больше среднего. В природе наиболее часто встречается пятнистое распределение, причем оно свойственно и наземным организмам, и обитателям водной толщи.
Наиболее важными показателями характеристиками популяции являются рождаемость и смертность. Рождаемость - это способность популяции к увеличению численности. Этот показатель используется и при анализе популяции человека, т.е. в демографии.
Максимальная рождаемость - это теоретический максимум скорости образования новых особей в идеальных условиях (когда отсутствуют лимитирующие факторы). Экологическая рождаемость или просто рождаемость обозначает увеличение численности популяции при фактических условиях среды. Эта величина непостоянна и варьирует в зависимости от размерного и возрастного состава популяции и физических условий среды. Обычно рождаемость выражают путем деления общего числа вновь появившихся особей на время, либо как число вновь появившихся особей в единицу времени на 1 особь в популяции (удельная рождаемость). Рассмотрим на примере различие между максимальной и реализованной, экологической рождаемостью: в популяции птиц отложено 510 яиц - около 15 на самку, это число соответствует максимальной рождаемости, оперилось всего только 265 птенцов - это значит, что экологическая рождаемость составляет около 8 птенцов на самку или 52 % от максимальной.
Смертность отражает гибель особей в популяции. Этот показатель противоположен рождаемость. Подобно рождаемости смертность выражает число особей, погибших за данный период. Экологическая смертность это смертность особей в данных условиях среды. Эта величина, как и экологическая рождаемость, не постоянна и изменяется в условиях среды.
Для экологов больший интерес, чем смертность, представляет выживаемость. Если число погибших особей выразить через С, то выживаемость В будет равна В = 1 - С. Полную картину смертности и рождаемости в популяции выражают статистическими таблицами выживания или демографическими таблицами, в которых указывают рождаемость, смертность, выживаемость, ожидаемую продолжительность жизни и т.д. для различных возрастных классов особей популяции. Графические зависимости, построенные на основе таблиц выживания, называют кривыми выживания. Типы кривых выживания представлены на рис. 8.
Различают три типа кривых выживания. Сильно выпуклая кривая свидетельствует о том, что смертность в популяции почти до конца жизненного цикла остается низкой. Такой тип кривой характерен для крупных животных и человека. Сильно вогнутая кривая - другой крайний вариант.
Такой вид кривой имеет место, когда смертность высока на ранних стадиях развития организмов (например, дуб). Кроме выпуклых и вогнутых типов кривых имеется промежуточный, когда удельная выживаемость для каждой возрастной группы одинакова. Видимо, в природе не существует популяций, у которых выживаемость постоянна (рис. 8), однако слегка выпуклые или вогнутые кривые встречаются часто.
Возрастная структура популяции является ее важной характеристикой, которая влияет и на рождаемость, и на смертность. Соотношение разных возрастных групп в популяции определяет ее способность к размножению в данный момент и показывает, чего можно ожидать в будущем.
Обычно в быстро растущих популяциях значительную долю составляют молодые особи, в популяциях, находящихся в стационарном состоянии, возрастное распределение более равномерно, а популяции, численность которой снижается, будет содержаться большая доля старых особей. Упрощенно в популяции можно выделить три экологические возрастные группы: предрепродуктивную, репродуктивную и пострепродуктивную.
Репродукция - воспроизведение особей. Длительность этих возрастов по отношению к общей продолжительности жизни варьирует у разных организмов.
У современного человека три этих "возраста" примерно одинаковы, хотя в странах, где население растет быстрыми темпами (Юго-Восточная Азия, Латинская Америка, Африка), более 40 % населения моложе 15 лет. Для многих животных и растений характерен очень длительный предрепродуктивный период, например, у некоторых насекомых предрепродуктивный период чрезвычайно длителен, репродуктивный - короткий, а пострепродуктивный период отсутствует (бабочки-поденки).
Популяции в естественных условиях изменчивы. Даже если сообщество и экосистема кажутся неизменными, их плотность, рождаемость, выживание, возрастная структура, скорость роста обычно изменяются по мере того, как вид подстраивается к сезонам, физическим факторам или другим видам.
Экологов интересует, с какой скоростью в популяциях происходят изменения. Так, если N представляет собой число организмов, а t - время, то N - изменение числа организмов, N /Nt - средняя скорость изменения числа организмов в расчете на 1 особь. Параметр N/Nt часто называют удельной скоростью роста и используют при сравнении популяций различной величины. Величина dN/dt будет представлять собой скорость изменения числа особей за единицу времени в некоторый момент, а dN /N dt - скорость изменения числа организмов в расчете на 1 особь в единицу времени в некоторый момент. Если окружающая среда не налагает никаких ограничений, то удельная скорость роста для данных микроклиматических условий становится постоянной и максимальной. Значение скорости роста в этих благоприятных условиях характеризует соответствующую возрастню структуру популяции и представляет собой единственный показатель наследственно обусловленной способности популяции к росту.
|

Его обозначают r:
r = dN/(N dt).
Путем интегрирования получаем экспоненциальную зависимость Nt в момент времени t:
Nt = N0 e rt,
где N0 - численность в начальный момент времени; е - основание натурального логарифма.
В логарифмической форме это уравнение удобно для проведения расчетов:
ln Nt = lnN0 + rt.
Показатель r - это фактически разность между удельной мгновенной скоростью рождаемости b и мгновенной скоростью смертности d и может быть выражен так: r = b - d. Максимальное значение r часто называют биотическим или репродуктивным потенциалом. Разность между максимальным r и скоростью роста в реальных условиях, часто называют сопротивлением среды, которая характеризует сумму всех лимитирующих факторов среды.
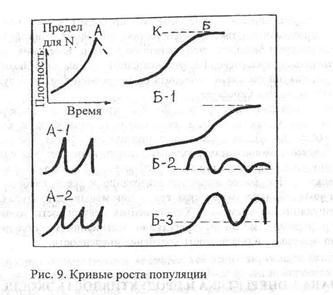
Характер увеличения численности популяции может быть различным, и в связи с этим выделяют различные типы роста популяции. По форме кривых роста можно выделить два основных типа: рост, описываемый J-образной и S-образной или сигмовидной кривой. Эти противоположные типы в соответствии с особенностями разных организмов и условий среды могут усложняться, модифицироваться или сочетаться различным образом (рис.9). При J-образной кривой плотность быстро возрастает по экспоненте, но затем, когда начинает действовать сопротивление среды или какой-то лимитирующий фактор, рост быстро прекращается. Этот тип роста может быть описан простым экспоненциальным уравнением: dN/dt = rN при заданном ограничении на величину N. При темпе роста, описываемом S-образной кривой, плотность вначале увеличивается медленно, а затем быстрее, приближаясь к логарифмической фазе, но вскоре под влиянием сопротивления среды рост постепенно замедляется. В конце концов, будет достигнуто равновесие, которое и сохраняется. Такой тип роста можно описать следующим уравнением:
dN/dt = r N [(К - N)/К].
Верхний предел увеличения численности популяции, соответствующий константе К, называется верхней асимптотой S-образной кривой. Ее называют пределом роста. При J-образном типе роста равновесие не достигается, напротив, плотность может колебаться в соответствии с ограничением ресурсов, физическими факторами и взаимодействием с другими популяциями. На рис. 9 показаны формы роста популяции: показаны J-образная кривая (А), S-образная кривая (Б) и некоторые их варианты. Варианты А-1 и А-2 представляют собой колебания, выявляемые при экспоненциальном росте, а Б-1, Б-2, Б-3 - некоторые, но далеко не все, возможные варианты роста, когда имеется разрыв во времени между увеличением плотности организмов, вызванным появлением новых особей и проявлением тормозящего влияния этого увеличения на рост популяции. Такой рост характерен для высших растений и животных.
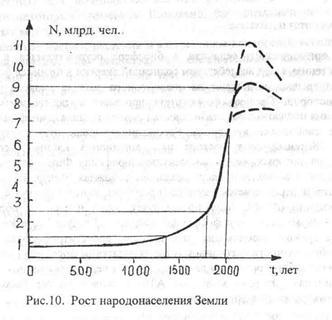
Если мы рассмотрим рост численности населения Земли, представленный на рис. 10, то увидим экспоненциальный характер роста, примерно с XVII-XVIII вв. нашей эры и по сегодняшнее время. Экспоненциальный рост свидетельствует о том, что пока численность населения Земли не достигла равновесия и продолжает расти. Ученые прогнозируют, что, если не произойдет резких перемен, такой рост популяции сохранится
и в XXI веке, пока численность не достигнет примерно 10 млрд человек. Такой прогноз основан на экстраполяции имеющихся данных по росту населения.
Однако углубляющиеся экологические проблемы ставят под сомнение возможность проживания такого количества людей в биосфере. Быстро истощающиеся ресурсы биосферы под давлением 5 млрд. населения, видимо, не смогут обеспечить проживание 10-миллиардного населения. Закономерности численности популяций позволяют говорить об изменении темпов будущего роста человеческой популяции в биосфере.
Когда популяция завершает рост и dN/dt в среднем за длительное время становится равным нулю, плотность ее проявляет тенденцию к флуктуациям (случайным отклонениям) относительно стационарного уровня. Часто флуктуации бывают обусловлены сезонными или годовыми изменениями доступности ресурсов. У некоторых популяций флуктуации столь регулярны, что их можно рассматривать как "циклические". Чем более высокоорганизованно и зрело сообщество живых
|
|
|
организмов и чем стабильнее физическая среда, тем меньше амплитуда флуктуаций плотности популяции во времени. Хозяйственная деятельность человека может нарушать природный механизм регуляции численности популяций, а следовательно, нарушать и цикличность колебаний численности.
2.5 Фотосинтез, хемосинтез и дыхание
Синтез органического вещества в биосфере осуществляется в основном зелеными растениями под воздействием солнечной энеpгии в процессе фотосинтеза. Растения непрерывно поглощают из атмосферного воздуха диоксид углерода и выделяют кислород. Процесс фотосинтеза протекает с поглощением энеpгии. Зеленые растения поддерживают стабильность газового состава атмосферы и наличие в атмосфере свободного кислорода. Рассмотрим упрощенную схему процесса фотосинтеза. Зеленый лист состоит из миллионов клеток, содержащих хлоропласты, в которых имеются молекулы хлорофилла. Фотон солнечного света взаимодействует с молекулой хлорофилла, высвобождая электрон одного из ее атомов. Электрон перемещается внутри хлоропласта, взаимодействуя с молекулой аденозиндифосфата (АДФ). АДФ представляет собой нуклеотид, образованный аденозином и двумя остатками фосфорной кислоты. В результате взаимодействия АДФ с электроном пpоисходит образование аденозитрифосфата (АТФ) - нуклеотида, образованного аденозином и тремя остатками фосфорной кислоты. Это вещество является универсальным аккумулятором и переносчиком энеpгии во всех живых организмах. Энергия молекулы АТФ в живой клетке расходуется на образование сахара из диоксида углерода и воды:
hv
6 H2О + 6 СО2 + АТФ → С6H12О6 + 6 О2 + АДФ
При этом АТФ превращается в АДФ, утрачивая часть энеpгии. Далее процесс повторяется. Описанный выше процесс фотосинтеза сопровождается сложными биохимическими реакциями, котоpые получили название темновых.
Ежегодно в ходе фотосинтеза в биосфере усваивается около 200 млрд т диоксида углерода и образуется 145 млрд т кислорода. Кроме зеленых pастений синтезировать органическое вещество могут некоторые бактерии. Источником углерода для них является диоксид углерода, но образования свободного кислорода при этом не пpоисходит. Бактерии, способные синтезировать сахар, носят название хемосинтетиков в отличие от pастений - фотосинтетиков. Например, при синтезе органического вещества серобактериями, химизм процесса состоит в следующем:
6 СО2 + 12 Н2 S → С6Н12О6 + 6 Н2О + 12 S
В продукции органического вещества хемосинтезирующие бактерии играют незначительную роль, но эти бактерии способны функционировать в условиях, неблагоприятных для большинства зеленых растений, а также участвовать в круговороте некоторых элементов. Возможно, бактериальный синтез полезен в загрязненных водах.
Большинство высших растений и многие виды водорослей используют только простые неорганические вещества, и, следовательно, являются полностью автотрофными. Некоторые водоросли, при том, что они синтезируют органическое вещество, нуждаются в каком-то одном органическом "ростовом" веществе, которое они сами не способны синтезировать, то есть частично они являются гетеротрофами. Организмы, занимающие промежуточное положение между автотрофами и гетеротрофами, называются ауксотрофными.
В глобальном масштабе эволюционно наиболее развитые формы жизни можно в основном разделить на автотрофов и гетеротрофов, причем для выживания последних необходим газообразный кислород.
Наряду с процессами синтеза органического вещества автотрофами имеет место процесс гетеротрофного разложения. Он примерно уравновешивает автотрофный метаболизм (обмен веществ). Если рассматривать разложение в широком смысле как любое биологическое окисление, дающее энергию, то с учетом потребности в кислороде можно выделить несколько типов разложения.
1. Аэробное дыхание, при котором окислителем служит газообразный кислород.
2. Анаэробное дыхание, протекающее без участия газообразного кислорода. Окислителем здесь служит какое-либо органическое или неорганическое вещество.
3. Брожение, в котором окислителем служит само органическое соединение.
Аэробное дыхание (тип 1) - процесс, обратный фотосинтезу: в нем синтезированное органическое вещество разлагается с образованием диоксида углерода и воды. Все высшие растения и животные получают энергию для поддержания жизнедеятельности и построения клеток с помощью этого процесса. Процесс дыхания может идти не до указанных конечных продуктов, и вместо воды и диоксида углерода могут образовываться органические соединения, которые используются организмами, осуществляющими разложение по типам 2 и 3. Бескислородное дыхание служит основой жизнедеятельности у сапрофагов - микроорганизмов, питающихся мертвой органикой (бактерии, дрожжи, плесневые грибы, простейшие), хотя как звено метаболизма оно может встречаться и в некоторых тканях высших животных. К общеизвестным организмам, использующим брожение (тип 3), относятся дрожжи, которые используются человеком, но, кроме того, дрожжи в изобилии встречаются в почве, где играют ключевую роль в разложении растительных остатков.
Разложение является результатом как биотических процессов, так и абиотических. Для некоторых экосистем, где физические условия таковы, что микроорганизмы не успевают разлагать органические остатки, агентом разложения может быть пожар, при котором растительные остатки сгорают, газы возвращаются в атмосферу, а минеральные вещества поступают в почву. Однако в основном окончательное разложение отмерших растений и животных осуществляют гетеротрофные микроорганизмы. Такое разложение есть результат процесса добывания пищи бактериями и грибами. Таким образом, разложение происходит, благодаря преобразованиям энергии внутри организмов и передаче ее между ними. Этот процесс абсолютно необходим для жизни. Прекратись он, все биогенные элементы вскоре оказались бы связанными в мертвых остатках, и стало бы невозможно продолжение жизни. Не все части растений и животных разрушаются с одинаковой скоростью. Жиры, сахара, белки разлагаются быстро, но растительная клетчатка, лигнин древесины, хитин, волосы и кости животных разлагаются очень медленно. Наиболее устойчивым продуктом разложения является гумус, который является обязательным компонентом всех экосистем.
В разложении органических остатков различают три стадии:
1) размельчение детрита в результате физического и биологического воздействия, сопровождаемое высвобождением растворенного органического вещества;
2) сравнительно быстрое образование гумуса и высвобо-ждение дополнительного количества растворимых органических веществ;
3) более медленная минерализация гумуса.
Обычно гумус имеет вид темного аморфного или коллоидного вещества. Физические свойства или химическое строение гумусовых веществ мало различается в географически удаленных или биологически различных экосистемах. С химической точки зрения гумусовые вещества представляют собой продукты конденсации ароматических соединений с продуктами распада белков и полисахаридов. Детрит, гумусовые вещества и другой органический материал, подвергшийся разложению, играют важную роль в образовании плодородных почв.
Таким образом, разложение органических остатков - длительный и сложный процесс, контролирующий несколько важных функций экосистемы. В результате этого процесса:
1) возвращаются в круговорот элементы питания;
2) образуются комплексы органических веществ с неорга-ническими;
3) с помощью микроорганизмов элементы питания и энер-гия возвращаются в экосистему;
4) производится пища для ряда организмов;
5) производятся вторичные метаболиты (вещества, образо-ванные в результате обмена веществ) ингибирующего, стимулирующего и регуляторного действия;
6) образуется почва;
7) поддерживается состав атмосферы, способствующий жизни аэробов, в том числе и человека.
2. 6 Трофические цепи и уровни
Живые обитатели Земли неодинаковы с точки зрения потребления вещества и энергии.
Синтезированное растениями органическое вещество передается в экосистеме от одних оpганизмов другим. Образуется цепь последовательной пеpедачи вещества и эквивалентной ему энеpгии, т. е. пищевая цепь или трофическая цепь.
Трофическая цепь - ряд видов или групп, каждое предыдущее звено, в котором служит пищей для следующего. Растения строят свой организм без посредников, самостоятельно, поэтому их называют самопитающимися или автотрофами. Они создают, продуцируют органическое вещество из неорганических и являются продуцентами. Далеко не все живые организмы могут строить собственное органическое вещество из минеральных, поэтому они используют в пищу то, что уже создано растениями - продуцентами. Такие организмы носят название гетеротрофов, т.е. питаемые другими. Второе название этих оpганизмов - консументы. Консументами являются все животные, часть микроорганизмов, паразитические и насекомоядные растения. Различают консументы первичные или первого порядка, к ним относятся растительноядные, и консументы вторичные или второго порядка, к ним относятся организмы, питающиеся животной пищей. Существуют консументы и более высоких порядков (третьего и четвертого).
Таким обpазом, продуценты и консументы обpазуют два первых звена трофической цепи. Особое звено трофической цепи составляют организмы, способные разлагать органические остатки до минеральных веществ, котоpые вновь могут быть усвоены растениями. Такие организмы носят название редуцентов и представлены бактериями, грибами, простейшими, беспозвоночными. Пищевые цепи могут быть простыми (осина - заяц - лиса) или более сложными (трава - насекомые - лягушки - змеи - хищные птицы), но очень длинными пищевые цепи быть не могут. Причина этого будет рассмотрена нами ниже. Число звеньев пищевых цепей составляет обычно от 3 до 5:
1) продуценты, то есть фотосинтезирующие растения;
2) первичные консументы, то есть растительноядные жи-вотные (фитофаги);
3) консументы второго порядка, то есть первичные хищ-ники, питающиеся растительноядными животными;
4) консументы третьего порядка - вторичные хищники, питающиеся первичными хищниками;
5) редуценты - микроорганизмы, бактерии, грибы, разла-гающие органическое вещество на минеральные составляющие.
Следует подчеркнуть особую роль редуцентов, разлагающих органические остатки и продуцентов, и консументов. Минеральные вещества, образующиеся при разложении, а также диоксид углерода, выделяющийся при дыхании редуцентов, вновь возвращаются к продуцентам. Особенно велика роль редуцентов в почвах.
Организмы, входящие в тpофические цепи, обpазуют тpофические уровни (рис. 12).
 Трофический уровень - совокупность оpганизмов, получающих преобразованную в пищу энергию Солнца и химических реакций через одинаковое число посредников трофической цепи: 1 уровень составляют продуценты, 2 - первичные консументы, 3 - вторичные консументы и т.д.
Трофический уровень - совокупность оpганизмов, получающих преобразованную в пищу энергию Солнца и химических реакций через одинаковое число посредников трофической цепи: 1 уровень составляют продуценты, 2 - первичные консументы, 3 - вторичные консументы и т.д.
Тpофические цепи в экосистемах являются и энергетическими цепями. Hапpимеp, консументы, потребляя органическое вещество продуцентов, получают энергию, которая расходуется на построение организма самого консумента, на его дыхание, теплоотдачу, движение. В экосистемах имеет место непрерывный поток энеpгии, заключающийся в передаче ее от одного пищевого уровня к другому.

 солнца СО2
солнца СО2
 | |||
 | |||
 Дыхание
Дыхание
Энергия








БЛОК ПРОДУЦЕНТОВ БЛОК КОНСУМЕНТОВ
Погибшие Отбросы, остатки
растения пищи, трупы животных
Минеральные
вещества БЛОК РЕДУЦЕНТОВ Дыхание
 |
. Схема пищевой цепи в биогеоценозе
При передаче энеpгии от уровня к уровню она рассеивается, теряется. Потери энеpгии компенсируются поступлением солнечной энеpгии.
2.7 Продуктивность экосистем
Каждая экосистема создает и расходует органическое вещество. Образование органического вещества в разных экосистемах протекает с разной скоростью. Hапpимеp, один га леса в год создает 7 т органического вещества, один га луга - 4 т. Это значит, что каждая экологическая система обладает определенной продуктивностью. Продуктивностью называется скорость образования биомассы органического вещества.
В процессе производства органического вещества выделяют 4 уровня продуктивности.
1. Валовая первичная продуктивность - это общая скорость фотосинтеза, включая и те органические вещества, которые были израсходованы на дыхание. Эту величину называют также "валовым фотосинтезом" или "общей ассимиляцией".
2. Чистая первичная продуктивность - скорость накопления органического вещества в растительных тканях за вычетом того органического вещества, которое использовалось при дыхании растений за исследуемый период. Эту величину называют также "наблюдаемым фотосинтезом" или "чистой ассимиляцией".
3. Чистая продуктивность сообщества - это скорость накопления органического вещества, не потребленного гетеротрофами (то есть чистая первичная продукция минус потребление гетеротрофами) обычно за вегетационный период или за год.
4. Вторичная продуктивность - это скорость накопления энергии на уровнях консументов.
Высокие скорости продуцирования наблюдаются в естественных и искусственных экосистемах там, где физические факторы благоприятны, и особенно при поступлении дополнительной энергии извне, уменьшающей собственные затраты на поддержание жизнедеятельности. Дополнительная энергия может поступать в разной форме: в тропическом дождевом лесу - в форме работы ветра и дождя, на возделываемом поле - в форме энергии топлива и работы, совершаемой человеком. В биологических сообществах процесс продуцирования непрерывен во времени.
Рассмотрим соотношение между поступлением солнечной энергии и продуктивностью, представленное. Как видно из таблицы, в экосистемах поглощается лишь около половины всей лучистой энергии солнца, в продукты фотосинтеза превращается самое большее 5 %.
|
|
|
|
|
Дата добавления: 2014-01-11; Просмотров: 2528; Нарушение авторских прав?; Мы поможем в написании вашей работы!