КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
BER, спаренная с репликацией
|
|
|
|
Тот же самый PCNA-связывающий мотив найден и в двух недавно описанных человеческих гликозилазах UNG2 и MYH1. Главными субстратами для этих гликозилаз служат некорректно встраивающиеся в процессе репликации урацил напротив аденина и аденин напротив 8-оксигуанина соответственно. UNG2 содержит PCNA-связывающий мотив в своей N-концевой части и является основной ДНК-урацил-гликозилазой человека. Напротив, N-конец UNG1, митохондриальной формы урацил-гликозилазы, содержит сигнал, указывающий на ее митохондриальную локализацию, но не PCNA-связывающий мотив. MYH является гомологом MutY E.coli, все ее формы несут PCNA-связывающий мотив в своем С-конце, вне зависимости от наличия у них сигнала митохондриальной локализации.
Предложено два объяснения возможного механизма, при котором эти две гликозилазы связываются с PCNA. Первое – обе эти гликозилазы могут преимущественно привлекать PCNА в район АР-сайта после выщепления неправильного основания, и таким образом направлять реакцию репарации по ее PCNA-зависимой ветви. Вторая возможность состоит в том, что UNG2 и MYH благодаря связыванию с PCNA могут ассоциироваться с «машиной репликации». Недавние исследования показали, что UNG2 может связываться с «машиной репликации» и через PCNA и через RPA (replication protein A, эукариотический гомолог белка SSB прокариот, состоящий из 3 субъединиц). Это больше подходит ко второму объяснению, но не отбрасывает и первого.
Урацил, являющийся субстратом UNG2, может попадать в ДНК двумя путями – при встраивании урацил-трифосфата во время репликации и дезаминировании уже встроенного цитозина. В первом случае, вновь встроенный урацил спаривается с аденином, и частота этого встраивания зависит от размера пула предшественника. Впрочем, надо помнить, что предшественник урацила совершенно «легально» постоянно присутствует в клетке и его уровень регулируется физиологическими механизмами. Во втором случае урацил оказывается спаренным с гуанином, причем 100–500 таких пар образуется в человеческой клетке ежедневно. UNG2 способна удалять урацил из обоих положений, две другие гликозилазы TDG и MED1 (MBD4) – только во втором случае (U/G). То есть урацил, встроившийся в процессе репликации может быть удален только UNG2, а урацил, появившийся в результате дезаминирования цитозина может быть убран тремя независимыми гликозилазами.
Похожая картина и с MYH1. Основной ее мишенью является аденин напротив 8-оксигуанина. Эта неправильная пара также образуется именно в процессе репликации ДНК. Здесь нужно отметить, что полимеразы ε и δ обычно вставляют именно аденин напротив 8-оксигуанина, а полимераза β – цитозин. Связывание MYH1 с PCNA может облегчать репарацию неправильного спаривания, возникшую в процессе репликации. Другая гликозилаза – OGG1 (FPG E.coli) способствует выщеплению 8-оксоG, который возникает при прямом окислении двунитевой ДНК, напротив цитозина, но не напротив аденина. OGG1 не несет PCNA-связывающего мотива и не нуждается в его помощи для выщепления 8-оксоG. Хотя пока нет точных экспериментальных подтверждений того, что MYH1 связывается с PCNA или с «машиной репликации», но аналогия с UNG2 напрашивается сама собой. Гликозилазы, несущие PCNA-связывающий мотив участвуют в репарации повреждений, возникающих именно в процессе репликации, в отличие от тех гликозилаз, которые подобные повреждения репарировать не способны.
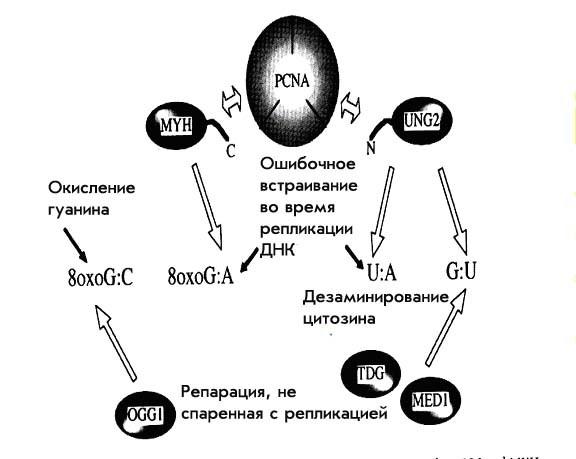
Рисунок 8. Схема репарации, спаренной с репликацией.
Таким образом две эти гликозилазы служат для специфической репарации, спаренной с репликацией, путем прямого связывания с репликационной машиной через PCNA. Схема этого процесса оитображена на рис. 8.
Остается нерешенным еще один вопрос – как эти две гликозилазы участвуют в репарации АР-сайтов. После действия UNG2 может включаться как PCNA-зависимый так и polβ зависимый процесс, так что тут все более-менее ясно. А вот MYH отличается тем, что не может использовать PCNA-зависимый путь, при котором синтез ведет polδ, так как polδ обязательно снова вставит аденин напротив 8-оксоG, и нарушение ДНК будет самовоспроизводиться в процессе репарации. Для репаративного синтеза может быть использован только polβ-зависимый путь, так как только polβ вставит напротив 8-оксоG цитозин
|
|
|
|
|
Дата добавления: 2014-11-06; Просмотров: 630; Нарушение авторских прав?; Мы поможем в написании вашей работы!