КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Структура генома
|
|
|
|
He секрет, что наука, как и любая область человеческой культуры, в огромной степени держится на мифах, т. е. устойчивых стереотипах сознания, когда спорные по своей сути и требующие доказательства положения принимаются массовым сознанием (в науке тоже есть массовое сознание) за аксиомы. Можно привести множество подобных мифов. В биологии одним из них является мнение, будто бы нам вполне ясен механизм наследственности и, что еще прискорбнее, будто бы нам ясен механизм развертывания наследственной информации в целостный организм.
Считается, что с открытием двуспиральной структуры ДНК и нуклеотидного кода здесь не осталось загадок. Но ведь геном человека состоит не из одной молекулы ДНК и даже не из серии молекул, расположенных внутри хромосом (к этому вопросу мы еще вернемся). Кроме того, существует морфогенетическая проблема - одна из главных проблем биологии в целом.
Суть ее в следующем: каким образом генетический код, записанный в том числе и в ядерных ДНК, развертывается в целостный организм? Действительно, нигде вы не найдете толкового объяснения, каким именно образом информация, записанная в хромосомах, реализуется в виде организованного ансамбля миллионов клеток, имеющих вполне определенную и устойчивую структуру. А ведь уже на ранних стадиях эмбрионального морфогенеза происходит специализация клеток, зародыш имеет определенную пространственную структуру и т. д.
С одной стороны, все клетки нашего организма - и нейроны мозга, и клетки печени, и клетки слизистой оболочки желудка - имеют одинаковый наследственный аппарат и возникли из одной единственной клетки - оплодотворенной материнской яйцеклетки, но насколько они внешне несхожи и насколько структурно несхожи образованные ими органы! Более того, специализированные клетки разных живых существ более структурно схожи между собой, чем клетки одного и того же существа, но составляющие разные ткани.
С другой стороны, неясно, каким образом яйцеклетка, а в дальнейшем клетки зародыша и взрослого организма, «узнают» структуру будущего слона или, скажем, бабочки, т. е. почему клетки изменяются нужным образом и в нужном месте и почему они единственно правильным образом располагаются в пространстве.
Впервые плодотворный подход к решению этой проблемы был предложен в 1920-е годы А. Г. Гурвичем, а в 1947 г. вышла в свет его книга «Теория биологического поля»[17], породившая, как это часто бывает с работами революционного характера, массу полуграмотных спекуляций - в ном случае, вокруг понятия «биополе» (сам термин и был предложен Гурвичем).
Еще на заре своей научной карьеры Гурвич открыл феномен, который назвал митогенетическим излучением (лучами Гурвича). Любопытно, что такая трактовка, как показали дальнейшие исследования, была в корне неверной, а вот сама концепция, созданная на ее основе, весьма продуктивной.
Лучи Гурвича возникают во время митоза и видны как своеобразные «протуберанцы», покрывающие клеточную мембрану. В интерфазе подобные явления не наблюдаются (рис. 17).
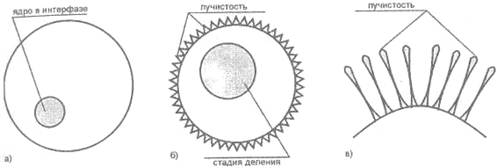
Рис. 17. Лучи Гурвичя
а) клетка в интерфазе;
б) «лучистость» - микроворсинки на поверхности клеточной мембраны;
в) структура микроворсинок
А. Г. Гурвич отождествил их с излучением электромагнитного характера, в том числе и в области видимого света. Несовершенство приборных методов исследования способствовало такой ошибке. В частности, исследования проф. А. Н. Мосолова (Медицинский ин-т, Новосибирск), к которым мы еще обратимся, показали, что митогенетическое излучение не светится в темноте и невоздействует на фотопленку.[18]
Однако именно Гурвич впервые в науке поставил вопрос о необходимости существования силового поля (биополя) сложной конфигурации, которое возникает в момент оплодотворения и существует в течение всей жизни организма, определяя структуру его среды. Делящиеся клетки подхватываются силовыми линиями, подобно тому, как это происходит с железными опилками в магнитном поле в известном школьном опыте. При этом геометрия поля реализуется в виде структуры организма, его органов и тканей, а проникая внутрь клеток, определяет их специализацию, т. е. «включение» тех или иных участков генома. Гурвич предполагал электромагнитную природу этого поля, а митогенетические лучи полагал его частным случаем.
Группа А. Н. Мосолова определила, что «лучистость Гурвича» на самом деле представляет собой густую сеть цитоплазменных выбросов типа микроворсинок (был предложен рабочий термин «пляшущие человечки»). Какова их природа? Удалось установить, что их порождает мощное акустическое поле. Далее удалось смоделировать данный процесс на вибростенде, подвергая механическим колебаниям поверхность вязкой жидкости (а именно такой и является цитоплазма и межклеточная среда).
Механизмы, определяющие функционирование и свойства акустических полей, достаточно сложны и разнообразны. Так, например, гидродинамический закон Бьеркнеса гласит, что два вибрирующих в газе или жидкости тела могут взаимно притягиваться или отталкиваться, в зависимости от их фазовых состояний.
Итак, существование мощных акустических полей в живом несомненно, их возможность структурировать среду таким образом, чтобы сделать деление клеток пространственно и хронарно не случайным, тоже (рис. 18). Но какие структуры связаны с этими вибрациями, и каким образом здесь задействован генетический аппарат?
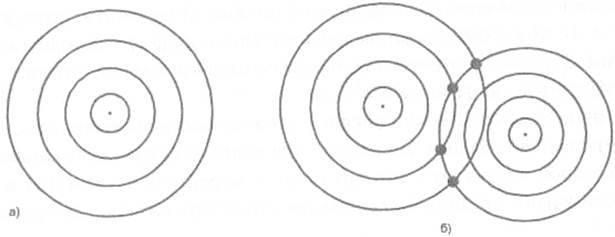
Рис. 18. Ноле точечных акустических источников
а) для одиночного источника;
б) для двух точечных источников.
Согласно распространенному заблуждению, генетический аппарат многоклеточных существует в двух формах: в виде клеточного ядра (в интерфазе) и в виде отдельных свободных хромосом (во время деления). Такое мнение сложилось потому, что в методах исследования внутриклеточных структур превалирует деструктивный подход. Тому способствуют фиксация, разрезание и раздавливание клеток при их микроскопировании. Таким образом, изучаются уже разрозненные обломки первоначальной структуры. Группа Мосолова поставила перед собой задачу изучения структуры именно живого, неповрежденного объекта. Были использованы методы фазовоконтрастной и интерференционной микроскопии. Удалось установить, что:
- источником микроскопического акустического поля является именно геном;
- таковой образует сложную пространственную структуру, по отношению к которой и отдельные хромосомы, и, тем более, хромосомное ДНК, представляют собой уже образование низшего порядка организации, отдельные детали общего механизма;
- автоколебательные физико-химические процессы, происходящие в недрах генетического аппарата, порождают автоколебательные акустические поля; таким образом, морфогенез - это, фактически, реализация сложной акустической голограммы.
В результате почти 30-летних исследований было установлено, что генетический аппарат - структурно единая, но пространственно динамическая система, в основе которой - многоуровневый сфероторои-дальный принцип организации. В интерфазе генетический аппарат принимает конфигурацию асимметричной сферы (это и есть клеточное ядро). Тела хромосом, укрепленные на внутренней мембране сферы ядра, располагаются меридианально. При этом все центромеры (кинетохоры) хромосом расположены у одного из полюсов, соединяясь в единую структуру - центромерное кольцо.
Определим некоторые используемые термины:
Интерфаза (лат. inter - «между» + греч. phasis - «проявление») -период жизненного цикла клетки между двумя митогенетическими делениями, в течение которого синтезируются вещества, необходимые Для существования и последующего деления клетки, а также возникают специальные структуры, в зависимости от ее функциональных особенностей; структурное поле проявлено слабо.
Митоз (греч. mitos - «нить») - основная форма клеточного деления, сущность которой заключается в удвоении числа хромосом и дальнейшем их равномерном распределении между дочерними клетками.
Морфогенез (греч. morphe - «вид», «форма» + греч. genes - «порождающий») - совокупность процессов развития оплодотворенной яйцеклетки, приводящих к формированию органов и систем зародыша (.морфогенез эмбриональный), а затем органов и систем организма.
Теломер (греч. telos — «конец» + греч. meros - «часть») – концевой участок хромосомы.
Хроматин (греч. chroma - «кожа», «цвет кожи») - вещество клеточного ядра, состоящее из ДНК, которая является носителем генетической информации, и небольшого количества РНК, а также белков; хорошо окрашивается основными красителями (отсюда название); в интерфазе, по представлениям официальной науки, рассеян в клеточном ядре, в период клеточного деления сконцентрирован в хромосомах. Исследования А. Н. Мосолова показывают, что и в интерфазе хроматин составляет вещество хромосом.
Хромосомы (греч. chroma - «кожа», «цвет кожи» + греч. soma -«тело») - структурный элемент клеточного ядра, содержащий ДНК, различимый, с точки зрения официальной науки, в виде образований определенного размера и формы только во время деления клетки, самоудвоение и закономерное распределение хромосом по дочерним клеткам обеспечивает передачу наследственной информации. По данным Мосолова, отдельно хромосомы не существуют - они образуют сложную пространственную форму, которая сохраняется и в интерфазе.
Центромера (лат. centreum - «центр круга» + греч. meros - «часть») - участок хромосомы в области ее первичной перетяжки.
Хвостовые участки хромосом (теломеры) сходятся у противоположного полюса ядра. Можно утверждать, что в естественном (неповрежденном) состоянии центромеры не существуют - это обрывки центромерного кольца. Точно так же не существуют и отдельные хромосомы. Такой подход позволяет разрешить и еще одну загадку биологии развития - механизм точного по количеству и качеству расхождения хромосом по дочерним клеткам.
Каждый вид живых существ имеет строго определенное количество хромосом. У человека их 46 (23 пары). Каждая хромосома имеет свою долю генетической информации и в этом смысле уникальна. При делении клетки происходит копирование генетического материала (репликация) и, следовательно, удвоение числа хромосом. Но каким образом хромосомы, если они никак не организованы, точно распределяются между дочерними клетками так, чтобы в каждой из этих клеток оказалось необходимое количество хромосом, и, при этом, их полный набор? Современная биология не знает ответа. Но если, как показывают исследования проф. Мосолова, все хромосомы представляют собой единую систему, то механизм их точного удвоения, а затем распределения между дочерними клетками под действием мощного акустического поля предстает простым и понятным (рис. 19).
Итак, организация генетического материала внутри ядра в пространственном отношении весьма сложна
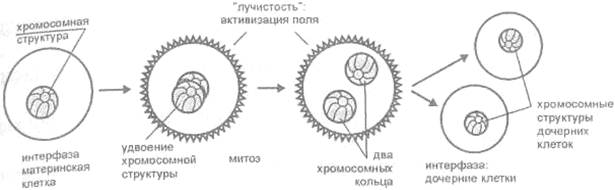
Рис. 19. Механизм удвоения и распределения
генетического материала ядра (по А. Н. Мосолову)
Но дело не только в этом. Мы должны признать, что, строго говоря, жизнь не ведает смерти. Действительно, рассмотрим внимательно ранние стадии развития многоклеточного живого существа, а именно человека (рис. 20).
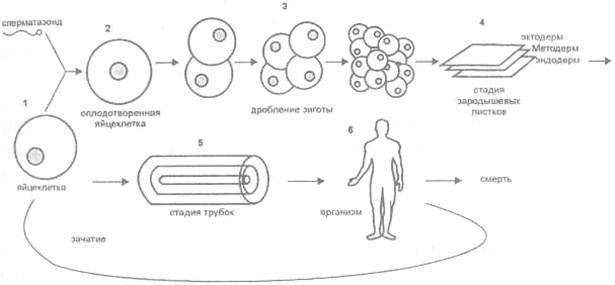
Рис. 20. Развитие человеческого существа
Первый этап связан с оплодотворением материнской клетки сперматозоидом. Далее происходит дробление зиготы - сперва вдвое, затем еще раз вдвое, затем - постоянное накопление клеточной массы без клеточной дифференциации. Следующий этап - перемещение цитологических масс, когда образуются три зародышевых листка - эктодерм, мезодерм и эндодерм. Далее происходит т. н. Первичный органогенез и формируется комплекс осевых органов: нервная трубка, кишечная трубка и хорда. Затем формируются собственно органы и, наконец, наступает момент рождения. Человек живет, умирает, но если он успевает породить потомство, то передает ему часть себя в виде цитоплазмы материнской клетки и прочих ее Неточных органелл.
Таким образом, геном передается не одним лишь клеточным (внутриядерным) ДНК, он состоит из:
- суммарного генетического материала отца и матери, сформированного сложным пространственным образом;
- акустического поля, возникающего как результат вибрации ядра, окруженного цитоплазмой и мембраной;
- клеточной цитоплазмы и других составляющих оплодотворенной яйцеклетки, чью преемственность, в принципе, мы можем проследить i вплоть до первого человека.
По мере зародышевого развития влияние наиболее глубинных структур размывается, а влияние собственно генной информации возрастает. Мы можем проследить следующие принципиальные этапы:
- этап непосредственно перед оплодотворением, когда вся совокупность будущего генома сосредоточена в двух половых клетках - отцовской и материнской;
- этап оплодотворенной материнской яйцеклетки, в которой проявлен весь геном и все его информационные носители (и, соответственно, составляющие) задействованы одновременно - и родительская цитоплазма, и акустическое поле, и генерирующий его и обладающий изначальной структурой ядерный генетический аппарат;
- этап дробления зиготы, когда в трансляции генома задейство- | ваны все три информационных носителя, клетки не дифференцированы и постепенно происходит падение значения родительской цитоплазмы; акустическое поле материнской клетки заменяется совокупным полем бластомеров, к концу этого этапа деление становится асинхронным;
- этап зародышевых листков, когда ситуация принципиально меняется: такой носитель, как родительская цитоплазма, уже не имеет никакого значения, клетки начинают дифференцироваться, таковой управляет конфигурация совокупного акустического поля, структурно весьма далекого от поля родительской клетки;
- этап первичного органогенеза, когда происходит дальнейшая дифференциация клеток, структура управляющего поля меняется, но оно еще носит, отчасти, единый общеорганизменный характер;
- этап зародышевого развития в форму, присущую человеческому существу, происходит пренатальная закладка всех органов и систем, единство акустического поля утрачивается;
- этап постпренатального роста и развития, созревания человеческого существа во взрослый организм, который начинается рождением, т. е. индивидуализацией человеческого существа, т. е. отделением его от материнского организма; он заканчивается половым созреванием;
- этап возмужания и старения; на этом этапе происходит «сброс» целостного генома в виде родительских половых клеток; трансляция собственного генома путем репликации ядерного генетического материала постепенно ухудшается - ошибки трансляции и приводят к смерти; _ этап умирания, деструкции генома в форму, не позволяющую поддерживать вертикальное единство человеческого существа (рис. 21).
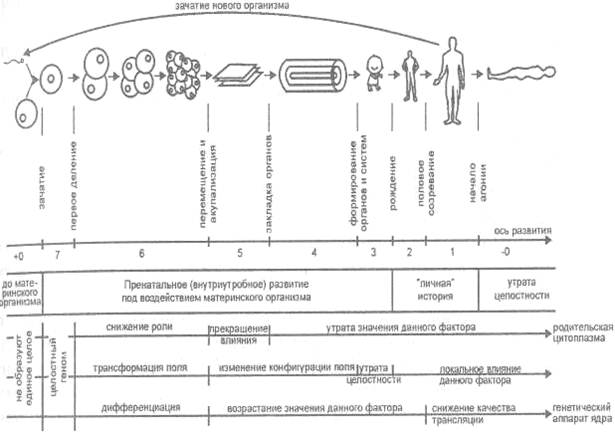
Рис. 21. Этапы формирования, развития, роста и умирания человеческого существа
Вновь мы видим, во-первых, выделенность девяти зон или уровней, из которых, во-вторых, лишь семь принадлежат собственно человеческому существу, а высшая (+0) лежит за пределами его пренатальной истории, низшая же (-0, смерть) знаменует собой полный распад единства. Все указанные зоны или уровни могут быть расположены вдоль оси развития, причем в двух «рядах» - генома и фенотипа, из которых второй - проявление первого.
Поскольку объем читаемой информации генома со временем уменьшается, определим генотип как читаемую часть генома и рассмотрим участки оси развития, учитывая, во-первых, ее двойственное проявление в виде серии генотипов и реализованных на их базе фенотипов, а, во-вторых, различие между зонами (уровнями оси) и участками читаемого генома, представляющими собой сумму низлежащих зон:
- Уровень (зона) 7 - зона Адама; ей соответствует целостный геном и полностью реализованный фенотип (назовем их иначе планетарными);
- уровень (зона) 6 - зона патриархов; ей соответствуют генотип и фенотип Homo;
- уровень (зона) 5 - зона пращуров; ей соответствуют генотип и фенотип этноса;
- уровень (зона) 4 - зона предков; ей соответствуют генотип и фенотип рода;
- уровень (зона) 3 - зона родителей; ей соответствуют генотип и фенотип семьи;
- уровень (зона) 2 - зона индивида; ей соответствуют генотип и фенотип индивида;
- уровень (зона) 1 - зона особи; ей соответствуют генотип и фенотип особи.
НЕ принадлежат собственно человеческому существу зоны:
- уровень (зона) + 0 - зона сянь («бессмертного»);
- уровень (зона) - 0 - зона смерти.
Уровни + 0, 7, 6 образуют участок ян, уровни с 5 по 3 - участок юань, уровни 2, 1,-0- участок инь.
Если рассматривать личную историю человеческого существа, то, начиная примерно с периода полового созревания и до существенного разрушения данных структур зоны особи, у него есть потенциальная возможность, включая путем соответствующей работы нечитаемые участки генома (ведь целостный геном всегда потенциально доступен), осуществить личную эволюцию из состояния инь - в состояние ян - и далее, к состоянию сянь, знаменующему начальную точку нового этапа развития Активного Сознания, в иной, «нечеловеческой» форме реализации. С этапами такой эволюции и связаны, главным образом, используемые нами (условные) названия зон и типов оси развития. Впрочем, как мы увидим в дальнейшем, они не произвольны, а отражают конкретные реалии, обусловленные структурными феноменами и фрактальностью организации, в том числе и «человеческого» мира.
На данном этапе изложения для пояснения сказанного мы можем изобразить развитие конкретного человека от базового «архетипа Адама» в виде следующей условной схемы (рис. 22).
Эволюция с подключением «спящих» информационных зон генома представляет собой, как мы видим, в определенном смысле, «возврат к Адаму»[19] или «обретение подобия».
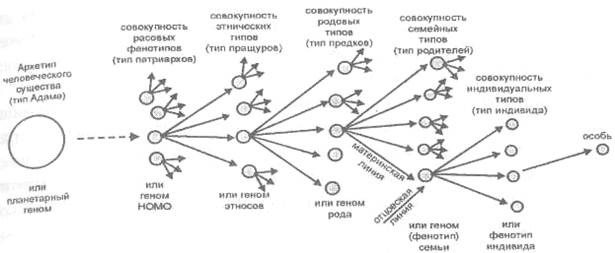
Рис. 22. Развертывание типа конкретного человека от базового архетипа Адама
Она идет, прежде всего, через восстановление целостного биополя, через зону юань и обретение «общечеловеческих качеств», не отменяющих, тем не менее, индивидуальности, которая лежит в основе витка «личной эволюции», в определенном смысле обратного витку «реализации начального фенотипа» (рис. 23).
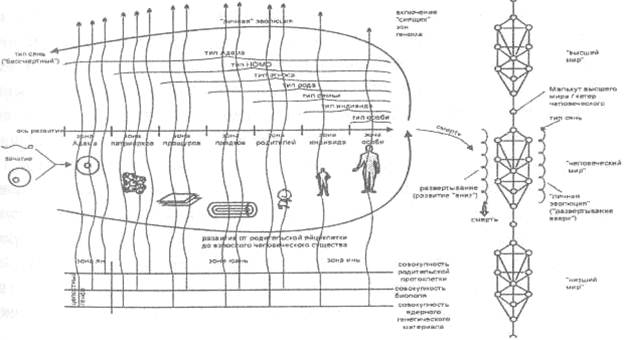
Рис. 23. Развитие от родительской яйцеклетки и личная эволюция
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 942; Нарушение авторских прав?; Мы поможем в написании вашей работы!