КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Строение сосудов хориоидеи. 1 страница
|
|
|
|
Артерии. Артерии не отличаются от артерий других локализаций и обладают средним мышечным слоем и адвентицией, содержащей коллагеновые и толстые эластические волокна (рис. 3.8.56). Мышечный слой от эндотелия отделен внутренней эластической мембраной. Волокна эластической мембраны переплетаются с волокнами базальной мембраны эндотелио-цитов [496, 959].
По мере уменьшения калибра артерии превращаются в артериолы. При этом исчезает сплошной мышечный слой стенки сосудов.
Сосуды и сосудистая оболочка глазного яблока

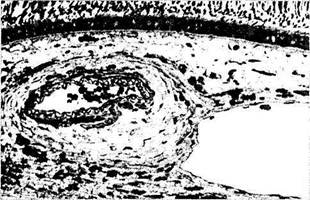
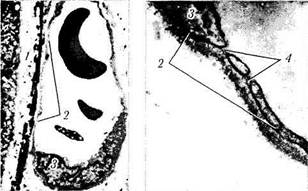
Рис. 3.8.56. Микроскопическое строение сосудов хорио-идеи:
а — артерия и вена хориоидеи крупного калибра (стенка артерии обладает толстым средним слоем и адвентицией); б, в — особенности ультраструктуры капиллярных сосудов хориоидеи (/—мембрана Бруха; 2 —эндотелиальная выстилка капилляра; 3 — ядро эндотелиальной клетки; 4 —дубликатура цитоплазмати-ческой мембраны с образованием «пор»)
Вены. Вены окружены периваскулярной оболочкой, вне которой располагается соединительная ткань. Просвет вен и венул выстлан эндотелием. Стенка содержит неравномерно распределенные гладкомышечные клетки в небольшом количестве. Диаметр самых больших вен равен 300 мкм, а самых маленьких, прека-пиллярных венул, — 10 мкм [154, 1028].
Капилляры. Капилляры хориокапиллярного слоя сосудистой оболочки имеют довольно большой просвет, позволяющий проходить нескольким эритроцитам. Выстланы они эндоте-лиальными клетками, снаружи которых лежат перициты (рис. 3.8.56, б, в). Количество перицитов на одну эндотелиальную клетку хориокапиллярного слоя довольно велико. Так, если в капиллярах сетчатки это соотношение равно 1:2, то в сосудистой оболочке— 1:6 [370, 708, 933]. Перицитов больше в фовеолярной области. Перициты относятся к сократительным клеткам и участвуют в регуляции кровоснабжения. Особенностью капилляров хориоидеи является то, что они фенестрированы, в результа-
те чего их стенка проходима для маленьких молекул, включая флюоросцеин и некоторые белки [ill, 1007]. Диаметр пор колеблется от 60 до 80 мкм. Закрыты они тонким слоем цитоплазмы, утолщенной в центральных участках (30 мкм). Фенестры располагаются в хориока-пиллярах со стороны, обращенной к мембране Бруха [496, 527] (рис. 3.8.57, в). Между эндо-телиальными клетками артериол выявляются типичные зоны замыкания.
Межклеточные контакты эндотелиальных клеток хориокапилляров особого типа. Близкие по строению контакты выявляются в синусоидах печени и венулах брыжейки [1208]. Выявляются неравномерно распределенные зоны замыкания и десмосомы, которые не полностью герметичны [875, 1028]. Между эндотелиальными клетками и перицитами существуют щелевые контакты [1028].
Иннервация сосудистой оболочки. Сосудистая оболочка иннервируется симпатическими и парасимпатическими волокнами, исходящими из ресничного, тройничного, крылонебного и верхнего шейного ганглиев (рис. 3.8.57). В глазное яблоко поступают они с ресничными нервами.
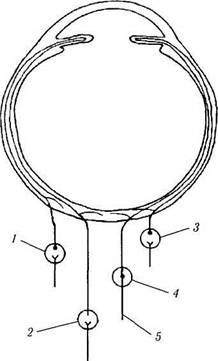
Рис. 3.8.57. Особенности вегетативной иннервации уве-ального тракта глаза:
/ — крылонебный ганглий; 2 —верхний шейный симпатический
ганглий; 3 — ресничный ганглий; 4 — тройничный ганглий; 5 —
пятый нерв
В строме сосудистой оболочки каждый нервный ствол содержит 50—100 аксонов, теряющих миелиновую оболочку при проникновении в нее, но сохраняющих шванновскую оболочку.
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 Постганглионарные волокна, исходящие из ресничного ганглия, остаются миелинизиро-ванными.
Постганглионарные волокна, исходящие из ресничного ганглия, остаются миелинизиро-ванными.
Сосуды надсосудистой пластинки и стромы сосудистой оболочки исключительно обильно снабжены как парасимпатическими, так и симпатическими нервными волокнами (рис. 3.8.58).
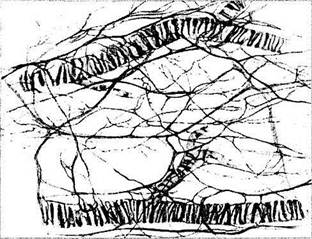
Рис. 3.8.58. Особенности распределения нервных волокон между сосудами сосудистой оболочки
Симпатические адренергические волокна, исходящие из шейных симпатических узлов, обладают сосудосуживающим действием.
Парасимпатическая иннервация сосудистой оболочки исходит от лицевого нерва (волокна, идущие из крылонебного ганглия), а также из глазодвигательного нерва (волокна, идущие из ресничного ганглия).
Последние исследования значительно расширили наши знания относительно особенностей иннервации сосудистой оболочки. У различных животных (крыса, кролик) и у человека артерии и артериолы сосудистой оболочки содержат большое количество нитрэргических и пептидэргических волокон, образующих густую сеть. Эти волокна приходят с лицевым нервом и проходят через крылонебный ганглий и не-миелинизированные парасимпатические ветви от ретроглазного сплетения [328, 1202]. У человека, кроме того, в строме сосудистой оболочки имеется особая сеть нитрэргических ган-глиозных клеток (положительны при выявлении НАДФ-диафоразы и нитроксидной синте-тазы), чьи нейроны связаны друг с другом и с периваскулярной сетью (рис. 3.8.59). Отмечено, что подобное сплетение определяется только у животных, имеющих фовеолу.
Ганглиозные клетки сконцентрированы в основном в височных и центральных областях сосудистой оболочки, по соседству с макуляр-ной областью. Общее количество ганглиозных клеток в сосудистой оболочке порядка 2000. Распределены они неравномерно. Наибольшее их количество обнаруживается с темпоральной
стороны и центрально. Клетки маленького диаметра (< 10 мкм) располагаются по периферии [328]. Диаметр ганглиозных клеток увеличивается с возрастом, возможно, из-за накопления в них липофусциновых гранул.
В нейронах выявлены нитрэргические трансмиттеры. Подобные нейротрансмиттеры обеспечивают расширение сосудов. Обнаруживаются они в периваскулярных нервах различных органов [116, 757, 787, 1081]. Этот медиатор вызывает также расслабление гладких мышц различных органов, например кишечника и трахеи [423], желчного пузыря [1059].
В некоторых органах типа сосудистой оболочки нитрэргические нейротрансмиттеры выявляются одновременно с пептидэргическими, также обладающими сосудорасширяющим действием [365, 614, 739, 1059]. Пептидэргичес-кие волокна [1118], вероятно, исходят из крылонебного ганглия и проходят в лицевом и большом каменистом нерве [1118]. Вероятно, что нитро- и пептидэргические нейротрансмиттеры обеспечивают вазодилятацию при стимуляции лицевого нерва.
Периваскулярное ганглиозное нервное сплетение расширяет сосуды сосудистой оболочки, возможно регулируя кровоток при изменении внутриартериального кровяного давления. Оно защищает сетчатку от повреждения тепловой энергией, выделяющейся при ее освещении. Flugel et al. [328] предложили, что ганглиозные клетки, расположенные у фовеолы, защищают от повреждающего действия света именно тот участок, где происходит наибольшая фокусировка света. Выявлено, что при освещении глаза существенно увеличивается кровоток в прилежащих к фовеоле участках сосудистой оболочки.

Рис. 3.8.59. Ганглиозная клетка сосудистой оболочки типичного строения, к которой подходит и контактирует нервное волокно:
/ — ганглиозная клетка; 2 —крупное ядрышко ганглиозной клетки; 3 — нервное волокно
Сосуды, и сосудистая оболочка глазного яблока
 Особенности кровообращения в сосудистой оболочке. Особенности кровообращения уве-ального тракта изучались интенсивно на протяжении многих лет как в эксперименте, так и в клинике. В 1975 г. Bill [114] суммировал имеющиеся данные и привел свою концепцию физиологии хориоидеи.
Особенности кровообращения в сосудистой оболочке. Особенности кровообращения уве-ального тракта изучались интенсивно на протяжении многих лет как в эксперименте, так и в клинике. В 1975 г. Bill [114] суммировал имеющиеся данные и привел свою концепцию физиологии хориоидеи.
У обезьян хориоидальный кровоток исключительно интенсивный, приблизительно в 20 раз выше, чем в сосудах сетчатой оболочки (радужка — 8 ± 1 мг/мл; ресничное тело — 81 ±6 мг/мл; сосудистая оболочка — 677 ± ±67 мг/мл; сетчатка — 34 ± 2 мг/мл). Поскольку интенсивность артериального кровотока столь высока, насыщенность кислородом венозной крови только на 3% ниже, чем насыщенность артериальной крови. И это несмотря на то, что кислород отдается наружной части сетчатой оболочки. Артериовенозные анастомозы играют небольшую роль в поддержании высокой насыщенности кислородом венозной крови. Предлагается, что высокий уровень увеаль-ного кровотока обеспечивает терморегуляцию внутриглазных оболочек, компенсируя снижение температуры в переднем отделе глаза и предотвращая перегревание сетчатки при ее освещении светом.
Регуляция кровотока. Механизмы регуляции кровотока в сетчатке и сосудистой оболочке существенно отличаются. Если в сетчатке преобладают механизмы ауторегуляции, то в хориоидее эти функции берут на себя симпатические нервные сплетения.
Интенсивность кровотока в сетчатке незначительно увеличивается при повышении концентрации рСО2 [347], а гипероксия вызывает небольшое сужение сосудов. При этом интенсивность кровотока снижается. Особенностью кровообращения сетчатки является и то, что на него не влияет изменение внутрисосудистого давления, что наблюдается, например, при изменении внутриглазного давления.
Кровообращение в хориоидее также усиливается при увеличении концентрации рСО2, но более значительно. При повышении парциального давления кислорода интенсивность кровотока практически не изменяется [112, 113].
Кровообращение хориоидеи не автономно, а регулируется нервными механизмами [248]. Ауторегуляция кровообращения выявлена только в сосудах ресничного тела и радужки. При стимуляции симпатической нервной системы наступает уменьшение просвета сосудов хориоидеи. При этом падает внутриглазное давление из-за уменьшения объема крови. Подобная реакция характерна для а-адренэргического типа иннервации [112, 113].
Сосуды хориоидеи находятся обычно в состоянии небольшого сокращения (сосудосуживающий тонус). Предполагают, что такое состояние защищает сетчатку от гиперперфузии сосудов, наблюдающейся при ряде заболеваний,
сопровождающихся повышением внутриартери-ального давления [119]. Вазомоторные терминалы заканчиваются в основном на артериолах и реже на артериях. Иннервируются также вены и венулы. Отсутствует иннервация хорио-капилляров [933]. На холинэргическую стимуляцию сосуды хориоидеи отвечают расширением просвета [1037].
В увеальном тракте выявлены также нит-ро- и пептидэргические волокна, обладающие сосудорасширяющим действием [933]. Подходят они к глазу по ходу лицевого нерва, образуя синапсы в крылонебном ганглии [328, 786, 1037, 1118].
Капилляры сосудистой оболочки и ресничных отростков напоминают таковые слизистой оболочки кишечника и почки. Исследование проницаемости этих капилляров выявило, что стенка пропускает большие молекулы. Дальнейшее продвижение молекул из сосудистой оболочки в сетчатку невозможно в результате наличия между пигментными клетками эпителия сетчатки плотных межклеточных контактов [222, 399, 1016]. Утечка белка из просвета капилляров сосудистой оболочки или ресничных отростков превышает подобную утечку в почках в пять раз, а в сердечной и скелетной мышцах в десять [120]. Благодаря такой высокой пропускной способности стенки сосудов, концентрация IgG в строме ресничных отростков и строме сосудистой оболочки составляет 60— 70% концентрации этого белка в плазме крови. Это свойство создает высокое осмотическое давление в ткани сосудистой оболочки (превышает давление сетчатки примерно на 15 мм ртутного столба). Разница в осмотическом давлении между сосудистой оболочкой и сетчаткой вызывает фильтрацию жидкости из сетчатки по направлению к сосудистой оболочке и является силой, которая придавливает сенсорную часть сетчатки к пигментному эпителию.
Наличие высокой проницаемости сосудов хориоидеи способствует транспорту в сетчатую оболочку витамина А, находящегося в макромо-лекулярном комплексе ретинол-связанного белка с преальбумином. Возможность выхода такой большой молекулы обеспечивается наличием фенестр. Высока пропускная способность сосудов и для низкомолекулярных веществ типа глюкозы. Причем она более чем в двадцать раз выше относительно сосудов мышцы сердца и в восемьдесят относительно сосудов скелетной мышцы. Это резко отличает сосуды хориоидеи от сосудов сетчатой оболочки.
Ишемия хориоидеи. На протяжении многих десятилетий непонятной оставалась причина развития ишемии хориоидеи при столь высокой насыщенности ее анастомозирующими сосудами. Причем участки ишемии хориоидеи строго очерчены [1198, 1199].
Механизмы развития локальной ишемии были непонятны и по следующим причинам:
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
1.  Кровоток в капиллярах хориоидеи один из
Кровоток в капиллярах хориоидеи один из
самых интенсивных — 800—1200/100 гр/мин.
2. К хориоидее направляется 85% всего
объема крови, направленного к глазному ябло
ку (к сетчатке только 4%).
3. Кровь в посткапиллярных кровеносных
сосудах хориоидеи столь же богата кислоро
дом, как и артериальная кровь [117, 293, 828].
4. Кровообращение в сосудах хориоидеи не
ауторегулируется, поскольку насыщение кро
ви углекислым газом минимальное [114, 117,
1196].
5. Максимальная стимуляция симпатичес
кой нервной системы приводит к уменьшению
объема кровообращения только на 60% [114,
117, 1196]; стенка капиллярных сосудов не гер
метична и проницаема для различных веществ,
включая белки.
Таким образом, имеющиеся анатомические и физиологические сведения не позволяли исследователям объяснить механизмы развития ишемии хориоидеи. Тем не менее, участки ишемии, а также инфаркта хориоидеи, не столь уж и редкое явление [77, 333, 515].
Были проведены многочисленные экспериментальные исследования, сводившиеся к введению в кровяное русло микрочастиц шаровидной формы [80, 210, 464, 893, 1198, 1199] или флюоресцеина [77, 257, 460, 515, 1108]. Но и при этом, объяснения этому явлению найдено не было. Лишь использование флюоресцеина позволило наблюдать наполнение участков хо-риокапиллярных сосудов флюоросцеином в виде секторов. Последовательность и площадь наполнения сосудов кровью четко соответствовала строению «хориокапиллярной дольки». Долька начинала наполняться кровью с центральных участков и лишь спустя несколько секунд кровь поступала к периферии. Поскольку строение «дольки» в различных участках уве-ального тракта различно (см. выше), различна и скорость кровенаполнения хориоидеи в различных участках.
Количественные и качественные характеристики кровообращения в «дольках» зависят от многих причин и, в первую очередь, от внутриглазного давления [257]. Установлено также, что венозная кровь отводится от каждой «дольки» в отдельности. При этом кровь соседних «долек» не смешивается. По всей видимости, такая система кровообращения предопределяет существование наиболее быстрого и короткого пути оттока венозной крови из хориоидеи. С другой стороны, возникает вероятность в определенных условиях возникновения ишемии хориоидеи на границе «долек». Наиболее часто ишемия наступает при окклюзии коротких задних ресничных артерий, а также сосудов глазницы [460, 470].
Развитие ишемии хориоидеи неблагоприятно влияет на строение и функции сетчатой оболочки. Исчезают фоторецепторы наружного ядер-
ного слоя, наступает миграция клеток пигментного эпителия в сетчатку.
Таким образом, на основании приведенных данных видна несостоятельность концепции о невозможности развития ишемии хориоидеи из-за большого количества анастомозов между сосудами.
Возрастные изменения сосудистой оболочки. В сосудистой оболочке глаза с возрастом уменьшается количество эластической ткани [1049], а также уменьшается толщина и самой сосудистой оболочки [868]. Вокруг крупных сосудов формируется широкая прослойка волокнистой ткани. При этом сосуды хориоидеи начинают напоминать сосуды радужки. В дополнение к описанному склерозу сосудистой стенки изменяется также число и калибр сосудов. Приведенные структурные изменения сопровождаются уменьшением скорости наполнения хориокапиллярного слоя, что показано при помощи флюоресцентной ангиографии. Появляются пятна гипофлюоресценции, хотя общая интенсивность свечения сохраняется независимо от возраста [525].
Регенерация увеального тракта. После повреждения любого участка увеального тракта наступает лишь заместительная регенерация. В эту область первоначально мигрируют клетки соединительной ткани (фибробласты), которые синтезируют межклеточное вещество и коллаген, выполняющие дефект. Затем наступает организация волокнистой ткани с образованием соединительнотканного рубца. Рубец, как правило, довольно интенсивно пигментирован, поскольку в нем скапливаются зерна меланина, высвободившиеся из поврежденных стромаль-ных меланоцитов. Существуют определенные различия в скорости заместительной регенерации радужной оболочки. Это связано с тем, что после ее повреждения (радиальные разрывы) края раны расходятся. В таких случаях рубцевания вообще не происходит.
Литература
1. Абрамов В. Г. Болезнь трансплантата рого
вицы.— Ярославль: Верх.-Волж. кн. изд-во, 1972. —
215 с.
2. Абрамов В. Г. К вопросу об иннервации рогови
цы // Офтальмол. журн.— 1959. — №6. — С. 358—
362.
3. Артемов А. В. Изменение дренажной зоны гла
за и сосудов, осуществляющих ее трофику в возраст
ном аспекте // Офтальмол. журн. — 1980. — № 7.—
С. 401—405.
4. Артемов А. В. Сравнительная характеристика
состояния тканей дренажной зоны глаза, сосудов ра
дужки и цилиарного тела у больных системными сосу
дистыми заболеваниями и простой глаукомой // Тез.
докл. Междунар. конф. офтальмологов городов-побра
тимов Одессы. — Одесса, 1981, —С. 181-182.
5. Артемов А. В. Состояние дренажной зоны и со
судов переднего отдела глаза у больных атероскле
розом, гипертонической болезнью, сахарным диабетом
Литература

 и открытоугольной глаукомой: Автореф. дис.... канд. мед. наук. — Одесса, 1982.— 20 с.
и открытоугольной глаукомой: Автореф. дис.... канд. мед. наук. — Одесса, 1982.— 20 с.
6. Беляев В. С. Склеропластика в лечении прогрес
сирующей миопии. — М.: Изд-во Ин-та дружбы наро
дов, 1977.— 229 с.
7. Боговягин В. Л., Франк Г. М. Субмикроскопичес
кая организация и функциональные особенности мюл-
леровских клеток сетчатки // Биофизика,— 1962.—
Т. 7. — № 1 — С. 42—50.
8. Вызов А. Л. Потенциалы в глиальных клетках
сетчатки // В кн.: Функции нейроглии.—Тбилиси:
Мецниереба, 1979. — С. 49—59.
9. Вит Ь. В., Дмитриев С. К- Гемофтальмический
барьер при травме глаза // Офтальм. журн. — 1997. —
№ 2.— С. 143.
10. Bum В. В., Мальцев Э.В. Особенности репара
ции повреждений эпителия роговицы и хрусталика у
животных, подвергшихся хроническому воздействию
малых доз ионизирующей радиации и интенсивному
световому облучению // Офтальм. журн. — 1998. —
№1. —С. 69—73.
М.Вит В. В., Мальцев Э. В., Павлюченко К-П. Влияние повышенной инсоляции и малых доз ионизирующей радиации на регенерацию эпителия хрусталика // Офтальм. журн. — 1997. — № 5. — С. 445—448.
12. Вит В. В., Юмашева А. А., Бабанина Ю. Д.
Послеоперационные осложнения циркляжа различ
ными материалами по данным экспериментальных
исследований // Офтальм. журн. — 1979. — № 4. —
С. 244—247.
13. Войно-Ясенецкий В. В., Думброва Н. Е. Ультра
структура многослойной волокнистой ткани, образую
щейся за десцеметовой оболочкой после ожога рого
вицы серной кислотой // Офтальм. журн.— 1971.—
№8.— С. 599—603.
14. Войно-Ясенецкий В. В. О природе и регенера-
ционных свойствах клеток стромы и эндотелия рого
вицы // В кн.: Материалы 3-й конференции по вопро
сам регенерации и клеточного размножения, 1962. —
С. 28-30.
15. Войно-Ясенецкий В. В. Патологический рост
эндотелия при экспериментальном ожоге роговицы
серной кислотой // Материалы научн. конф., посвя
щенной 90-летию со дня рождения В. П. Филатова,
Киев. 1965 —С. 19—20.
16. Войно-Ясенецкий В. В. Процесс приживления
роговичного трансплантата при внутричерепном по
вреждении тройничного нерва // Офтальм. журн. —
1959. — № 3. — С. 170—176.
17. Войно-Ясенецкий В. В. Метаплазия тканей гла
за при осложненном раневом процессе // В кн.: Усло
вия регенерации органов и тканей у животных. — М.,
1965 — С. 45—49.
18. Войно-Ясенецкий В. В. Разрастание и изменчи
вость тканей глаза при его заболеваниях и травмах. —
К.: Вища школа, 1979. — 224 с.
19. Калинина А. В. Глиальные клетки сетчатки ля
гушки Rana ridibunda Pall // Арх. анат., гист. и эмб-
риол. — 1983. — Т. 84. — № 4. — С. 33—38.
20. Кашинцева Л. Т. Глаукома у больных сахарным
диабетом // Автореф. дис.... д-ра мед. наук. — М.,
1972.— С. 30.
21. Красновид Т. А., Вит В. В. Повреждение и вос
становление функции клеток заднего эпителия роговой
оболочки после экстракции катаракты // Офтальмол.
журн. — 1995. — № 3. — С. 158
22. Мальцев Э. В. Хрусталик. — М.: Медицина,
1988.— С. 190.
23. Мальцев Э. В., Павлюченко К. П. Биологичес
кие особенности и заболевания хрусталика. — Одесса:
Астропринт, 2002. — 445 с.
24. Назаренко Н.И., Вит В. В., Бабанина Ю. Д.
Динамика морфологических изменений оболочек гла
за и прочности склеры после диатермокоагуляции //
Офтальм. журн. — 1981. — № 8. — С. 498—501.
25. Певзнер Л. 3. Биохимические особенности гли
альных клеток как основа для участия нейроглии в
специфической активности нейронов // В кн.: Функции
нейроглии. — Тбилиси: Мецниерба, 1979. — С. 251 —
265.
26. Певзнер Л. 3. Функциональная биохимия нейро
глии.—Л.: Наука, 1972.— 200 с.
27. Полунин Г. С, Макаров И., Шеремет Н. Осо
бенности клинического течения отдельных видов луче
вых катаракт // Вестн. офтальмол. — 1998. — № 5. —
С. 32—35.
28. Пучкшська Н. О. До питания про морфолопю
Hepeie i нервових закшчень poroeoi оболонки // Ме-
дичн. журн. — 1947. — № 16. — С. 340—356.
29. Пучывська Н. О. Морфолопчш особливост1 нер-
bjb ештелш роговоТ оболонки // В кн.: 36., присвяч.
Пям'ят1 О. В. Леонтовича (1869—1943). — К., 1948. —
С. 142—149.
30. Пучковская Н. А., Войно-Ясенецкий В. В. Вто
ричные дистрофические и структурные изменения
в переднем отделе глаза. — М.: Медицина, 1985.—
С. 192.
31. Розенфельд И. А. Флюорометрия в офтальмоло
гии. Обзор литературы // МРЖ. Офтальмология. —
1987. — № 4. —Т. 448.— С. 25—30.
32. Судакевич Д. И. Архитектоника системы внут
риглазного кровоснабжения. — М.: Медицина, 1971. —
С. 111.
33. Тринчук В. В., Мальцев Э. В., Расина Д. Г.,
Бормусова Э. А. Гистологическая и гистохимическая
характеристика ожогов роговицы различной степени
тяжести // Тез. докл. 1-го укр. съезда анатомов, гис
тологов и топографоанатомов.—Винница, 1980.—
С. 127—128.
34. Тринчук В. В., Мальцев Э. В., Расина Д. Г.,
Бормусова Э. А. Диэлектрические и патогистологичес-
кие параллели моделированных имических ожогов ро
говой оболочки глаза // Офтальмол. журн. — 1985. —
№2.— С. 115—118.
35. Федоров С. #., Егорова Э. В. Хирургическое
лечение травматических катаракт с интраокулярной
коррекцией. — М.: Медицина, 1985. — 327 с.
36. Федоров С.Н., Ронкина Т. И., Явишева Т. М.
Эндотелий роговицы человека.—М., 1993. — С. 126.
37. Шибкова С. А. О ганглиозных клетках сетчатки
лягушки // Арх. анатомии, гистологии и эмбриоло
гии. — 1 970. — Т. 9. — № 11. — С. 72—77.
38. Шибкова С. А. О ганглиозных клетках сетчат
ки селахий // Арх. анатомии, гистологии и эмбриоло
гии. — 1971. — Т. 60. — №3. — С. 21—28.
39. Школьник-Яррос Е. Г., Калинина А. В. Нейро
ны сетчатки. — М.: Наука, 1986. — 205 с.
AO.Acharya S., Rodriguez /., Moreira Т. SPACR a novel interphotoreceptor matrix glycoprotein in human retina that interacts with hyaluronan // J Biol Chem. — 1998.— Vol. 273.— P. 31599—31606.
41. Acott T.S., Samples J. R., Bradley J. M. Trabe-
cular re-population by anterior trabecular meshwork
cells after laser trabeculoplasty // Am J Ophthalmol. —
1989.— Vol. 1. —P. 107—112.
42. Adler A. J., Martin K. /. Retinol-bilding in bovine
interphotoreceptor matrix // Biochem Biophys Res Com-
mun—1982.— Vol. 108.— P. 1601 — 1608.
AZ.Ahnelt P.K., Keri C, Kolb H. Identification of pedicles of putative blue sensitive cones in human and primate retina // J Comp Neurol. — 1990. —Vol. 293 — P. 39—53.
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
44.  Ahnelt P. К., Kolb И., Pflug R. Identification of
Ahnelt P. К., Kolb И., Pflug R. Identification of
a.subtype of cone photoreceptor likely to be blue sensi
tive in the human retina // J Camp Neurol— 1987. —
Vol. 255 (18).— P. 34—40.
45. Ahnelt P., Kolb H. Horizontal cells and cone
photoreceptors in human retina: A Golgi-electron micro
scopic study of spectral connectivity // J Compar Neu
rology. — 1994. — Vol. 343. — P. 406—427.
46. Ahnelt P., Kolb H. Horizontal cells and cone
photoreceptors in primate retina: A Golgi-light micro
scope study of spectral connectivety // J Comp Neu
rol — 1994. — Vol. 343. — P. 387—405.
M.Aitken D., Friend J., Thoft R. A. Corneal re-epi-thelialization from the conjunctiva // Invest Ophthalmol Vis Sci. — 1988. — Vol. 29. — P. 224—231.
48. Aizawa K. The depth of the normal anterior
chamber // Acta Soc Ophthalmol.—1958. —Vol. 62.—
P. 2283—2289.
49. Al-Aswad L., Adorante J.S., Erickson К. E. Ef
fects of cell volume regulators on outflow facility in calf
and human eyes in vitro (Abstract) // Invest Ophthal
mol Vis Sci. — 1995. — Vol. 36. — P. 3331—3338.
50. Albona /., Purslowb P.P., Karwatowskic W.S.S.,
Eastyd D. L. Age related compliance of the lamina cri-
brosa in human eyes // Br J Ophthalmol. — 2000. —
Vol. 84.— P. 318—323.
51. Albona J. An investigation into the age-related
changes in the extracellular matrix of the human lamina
cribrosa // PhD thesis. — Bristol: University of Bristol.,
1995.
52. Alcala I., Maisel H. Biochemistry of lens plasma
membranes and cytoskeleton // In The Ocular Lens:
Structure, Function and Pathology / Ed. H. Maisel, Mar
cel Dekker. — New York, 1985. — 169 p.
53. Allansmith M. R., Kajiyama G., Abelson M. B.
Plasma cell content of main and accessory lacrimal
glands and conjunctiva // Am J Ophthalmol. — 1976. —
Vol. 82 —P. 819—825.
54. Allen D.P., Low P.S., Dola A. Band 3 and an-
kyrin homologues are present in the eye lens: Evidence
for all major erythrocyte membrane components in same
non-erythroid cell // Biochem Biophys Res Commun. —
1987.—Vol. 149—P. 266—273.
bb.Allsopp R.C., Vaziri #., Patterson C, Goldstein S., Younglai E. V., Futcher A. B. Telomere length predicts replicative capacity of human fibroblasts // Proc Natl Acad Sci. — 1992. — Vol. 89. — P. 10114—10118.
56. Almegard В., Andersson S. E. Outflow facility
in the monkey eye: Effects of calcitonin gene-relat
ed peptide, cholecystokinin, galanin, substance P and
capsaicin // Exp Eye Res. — 1990. — Vol. 51 —
P. 685—692.
57. Alvarado J. A., Van Horn С Muscle cell types of
the cat inferior oblique // In Lennerstrand G., Bach-y-
Rita P. (eds): Basic Mechanisms of Ocular Motility. —
Oxford Pergamon Press, 1975. — P. 15—45.
58. Alvarado J. A., Yun A.]., Murphy C.G. Juxta-
canalicular tissue in primary open angle glaucoma and
in nonglaucomatous normals // Arch Ophthalmol. —
1986.— Vol. 104.— P. 1517—1525.
59. Alvarado /., Murphy C, luster R. Age-related
changes in the basement membrane of the human cor
neal epithelium // Invest Ophthalmol Vis Sci.— 1983.—
Vol. 24.— P. 1015—1021.
60. Alvarado /., Murphy C, luster R. Trabecular
meshwork cellularity in primary open angle glaucoma
and nonglaucomatous normals // Ophthalmology. —
1984.—Vol. 91. —P. 564—572.
|
|
|
|
|
Дата добавления: 2014-11-18; Просмотров: 670; Нарушение авторских прав?; Мы поможем в написании вашей работы!