КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
РНК и её роль в сохранении и реализации наследственной информации
|
|
|
|
Химический состав РНК. Атипичные азотистые основания РНК. Макромолекулярная организация РНК. Типы РНК и их биологические функции.
Химический состав, атипичные азотистые основания и макромолекулярная организация РНК рассмотрены ранее (см. 1.2.2). Рибонуклеиновые кислоты (РНК), присутствующие в клетках как про-, так и эукариот, бывают трех основных типов: информационная (матричная, мРНК), транспортная (тРНК) и рибосомная (рРНК). В ядре клеток эукариот содержится РНК четвертого типа - гетерогенная ядерная РНК (гяРНК). У некоторых вирусов РНК служит носителем генетической информации. мРНК является копией (транскриптом) соответствующей ДНК. Этот транскрипт служит матрицей для синтеза белка. Каждые три последовательных основания мРНК (называемые кодоном) детерминируют один аминокислотный остаток. Молекулы тРНК переносят специфические аминокислотные остатки к определенному участку мРНК в ходе синтеза белка. В настоящее время структура тРНК хорошо известна. Молекулы рРНК встречаются в различных формах и образуют в комплексе с белками рибосому - сложную органеллу, в которой происходит синтез белка.
ТРАНСПОРТНАЯ РНК (тРНК) узнает соответствующий кодон в мРНК и переносит нужную аминокислоту к растущей полипептидной цепи. Узнавание кодона в мРНК осуществляется с помощью трех последовательных оснований в тРНК, называемых антикодоном. Между азотистыми основаниями кодона и антикодона образуются уотсон-криковские водородные связи при условии, что полинуклеотидные цепи антипараллельны. Аминокислотный остаток может присоединяться к З'-концу молекулы тРНК. Специфичность такой системы переноса обеспечивается тем, что имеется, по крайней мере, одна тРНК для каждой аминокислоты. Так, тРНК для Рhе обозначается тPHKPhe.
В одной эукариотической клетке более ста различных молекул мРНК, каждая из которых кодирует одну или несколько полипептидных цепей.
Молекулы тРНК состоят примерно из 75 нуклеотидов и начинаются с фосфорилированного 5'-конца. Первым основанием обычно является гуанин. На З'-конце всегда присутствуют три основания -ССА и концевая ОН-группа. В структуре тРНК, кроме обычных, часто присутствуют минорные основания. Исходя из максимального числа уотсон-криковских пар оснований при данной нуклеотидной последовательности, структуру молекулы тРНК принято представлять в виде «клеверного листа».
Участки, в которых посредством Н-связей образовались пары оснований, называются стеблями, а одноцепочечные участки — петлями. Все известные тРНК образуют «клеверный лист» с четырьмя стеблями (акцепторным, D, антикодоновым и Т) и тремя петлями (D, антикодоновой и Т). Некоторые тРНК имеют дополнительные петли и/или стебли (например, вариабельная петля
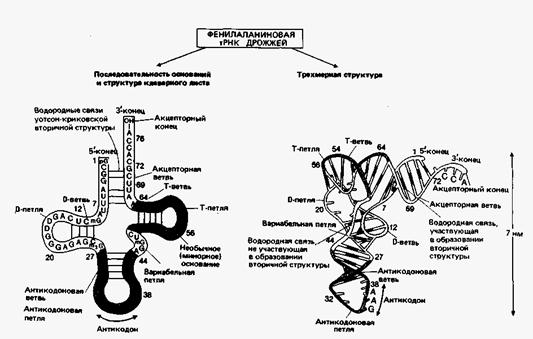
Рис.17. Строение дрожжевой тРНК
дрожжевой фенилаланиновой тРНК). Каждый стебель состоит из двух антипараллельных цепей, образующих правую двойную спираль, известную как А-форма РНК. Эта форма содержит 11 пар оснований на виток, шаг спирали равен 3.1 нм. Плоскости оснований составляют  около 20° с нормалью к оси двойной спирали. А-форма РНК близка к аналогичной форме ДНК. РНК не способна менять конформацию и переходить в В-форму, что обусловлено наличием 2'-ОН группы в рибозе, которой нет в дезоксирибозе. Пространственная (третичная) структура молекулы тРНК напоминает по форме букву Г. «Перекладину» этой буквы образует спираль из акцепторного и Т-стеблей, а антикодоновый и D-стебли формируют «ножку» (рис.17). В каждой части содержится примерно 10 пар оснований.
около 20° с нормалью к оси двойной спирали. А-форма РНК близка к аналогичной форме ДНК. РНК не способна менять конформацию и переходить в В-форму, что обусловлено наличием 2'-ОН группы в рибозе, которой нет в дезоксирибозе. Пространственная (третичная) структура молекулы тРНК напоминает по форме букву Г. «Перекладину» этой буквы образует спираль из акцепторного и Т-стеблей, а антикодоновый и D-стебли формируют «ножку» (рис.17). В каждой части содержится примерно 10 пар оснований.
рРНК играют важную роль в структуре и биосинтетической функции рибосом — органелл клетки, состоящих из двух субчастиц, на которых происходит синтез белка. Каждая такая частица образована молекулами протеинов и рРНК. Коэффициент седиментации рибосом прокариот составляет примерно 70S, а для эукариот он равен 80S. Своими собственными рибосомами обладают и митохондрии эукариотических клеток. Их коэффициент седиментации равен 70S, и они во многом сходны с рибосомами прокариот. 70S-рибосома прокариот состоит из 50S- и 30S-субчастиц, а SOS-рибосома эукариот — из 60S- и 40S-субчастиц. Эти субчастицы, в свою очередь, могут диссоциировать на составные части — белок и рРНК. Весовое соотношение между ними для про- и эукариот составляет 2:1 и 1:1 соответственно. Большинство рибосомных белков содержит в боковых цепях остатки Arg+ и Lys+. Эти положительно заряженные группы взаимодействуют с отрицательно заряженными фосфатными остатками молекул рРНК, за счет чего стабилизируется комплекс протеин-НК. Для некоторых молекул рРНК установлена нуклеотидная последовательность, в которой обнаружены участки спаривания оснований, образующие вторичную структуру, подобную той, что имеет место в стеблях молекулы тРНК.
Матричный синтез РНК осуществляется с помощью ДНК-зависимых РНК-полимераз, для которых характерна мультисубъединичная структура. В строении этих Ф у прокариот и эукариот имеются существенные различия. Так, в эукариотических клетках представлены три вида РНК-полимераз. Их принято обозначать римскими цифрами либо заглавными буквами латинского алфавита. РНК-полимераза I (А) локализована в ядрышке, где она катализирует синтез предшественников больших рибосомных РНК; РНК-полимераза II (В) находится в нуклеоплазме и участвует в синтезе первичного транскрипта мРНК; полимераза III (С) также локализована в нуклеоплазме и катализирует синтез тРНК и 50S-pPHK.
Инициация транскрипции происходит на строго определенном участке матрицы, расположенном после промотора. Последний представляет собой участок молекулы ДНК, содержащий несколько десятков пар оснований и находящийся вне пределов области, программирующей синтез РНК. В свою очередь синтез РНК всегда начинается с оснований А или G в «+»-цепи ДНК, а участок связывания Ф расположен выше точки инициации примерно на 10 оснований.
Элонгация цепи РНК наступает после присоединения нескольких нуклеозидов. В этот момент РНК-полимераза претерпевает структурные изменения: у прокариот от Ф отделяется σ-субъединица, а оставшийся кор-фермент — комплекс (а2ββ') субъединиц — катализирует дальнейшее удлинение цепи РНК. Движущийся вдоль ДНК кор-фермент действует подобно застежке-молнии и раскрывает двойную спираль, которая замыкается позади Ф по мере того, как соответствующие основания РНК спариваются с основаниями ДНК в «минус» — цепи.
Терминация цепи РНК происходит на специфических участках ДНК — терминаторах. РНК чаще всего образуются в виде предшественников, которые, уже без участия матричных систем, превращаются в зрелые молекулы. Всю совокупность происходящих при этом превращений как с РНК, так и с белками называют процессингом. Например, молекулы мРНК образуются из больших по размеру предшественников, называемых гетероядерной РНК (гяРНК). Для образования зрелой мРНК из этих молекул ферментативным путем удаляются лишние концевые олигонуклеотиды (по 5'- и З'-концам), а также имеет место «монтаж» (сплайсинг) — удаление крупных вставочных последовательностей, соответствующих интронам (некодирующим участкам) ДНК, с одновременным воссоединением концов соседних экзонов (кодирующих участков) в единую цепь. Наконец, многие РНК, в первую очередь тРНК, подвергаются многочисленным химическим превращениям, приводящим к образованию в составе зрелой молекулы широкого спектра минорных компонентов, таких как дигидроуридин, 4-тиоуридин, инозин и др. Следует подчеркнуть, что Ф, участвующие в транскрипции, являются мишенями для ряда БАВ, в частности антибиотиков и токсинов. Например, токсин бледной поганки - α-аманитин - блокирует РНК-полимеразу II эукариот, что приводит к прекращению синтеза новых молекул мРНК и многих жизненно важных белков.
Длительное время биологи просто не обращали особого внимания на короткие отрезки клеточной рибонуклеиновой кислоты (РНК), полагая, что их роль в клетке не слишком значительна. Гораздо больший интерес привлекали другие типы РНК, а именно информационные и рибосомалъные. Оба этих класса - очень длинные молекулы, содержащие до 100 000 нуклеотидов. Первые (информационные, которые часто называют также матричными РНК, или мРНК) переносят генетическую информацию с хромосом (ДНК) в специальные органеллы - «агрегаты» для синтеза белков - рибосомы. Второй класс - рибосомальная РНК - является одновременно и строительным материалом, и важнейшей рабочей частью рибосом.
В последние годы была установлена роль коротких отрезков РНК. Понятно, что с первого взгляда малые РНК, состоящие всего из нескольких десятков нуклеотидов, могли показаться остатками от своих «больших братьев». И даже несмотря на то, что роль отдельных малых молекул РНК в процессах превращения информационных РНК (сплайсинге), а также при упаковке нитей нуклеиновых кислот, была доказана ранее, истинным «хитом» в биологии малые РНК стали только лишь с открытием своей способности подавлять экспрессию генов у животных.
В нормально работающей клетке каждый ген выполняет собственную, строго определенную функцию, например, отвечает за выработку белка, мРНК, или за взаимодействие с другими регуляторными белками. При этом говорят о нормальной экспрессии (от лат. expressus - выразительный, явный) гена в клетке. Если же количество продукта данного гена (например, белка) снижается, то говорят о снижении экспрессии данного гена. Эффект «гашения» экспрессии определенных генов малыми РНК получил название РНК - интерференции, а молекулы, вызывающие его, назвали siRNA (small interfering Ribonucleic Acids - малые интерферирующие рибонуклеиновые кислоты). С открытием siRNA - интерференции стало ясно, что этот феномен может иметь огромное практическое значение.
В класс малых РНК включают молекулы, содержащие от 20 до 300 нуклеотидов. За эффект РНК - интерференции отвечают самые короткие из них - siRNA, состоящие всего из 21-28 (у млекопитающих из 21-23) нуклеотидов. Особенностью этих молекул является то, что они, в отличие от большинства других клеточных РНК, состоящих всего из одной цепи нуклеотидов, являются двунитчатыми. Нуклеотиды с противоположных нитей (цепей) siRNA спариваются друг с другом по тем же законам комплементарности, которые формируют двунитчатые цепи ДНК в хромосомах. Кроме того, по краям каждой из цепей siRNA всегда остается два неспаренных нуклеотида. Как siRNA появляются в клетке? Очевидно, в клетке должен существовать некий молекулярный механизм, который обеспечивал бы синтез siRNA, их накопление в клетке и позволял бы им выключать гены. Ученым удалось выявить систему ферментов, которая во многом схожа у всех многоклеточных и некоторых одноклеточных организмов.
Если молекула siRNA по тем или иным причинам (например, по воле исследователя) появляется в клетке, ее сразу же «берет в оборот» специальная клеточная система белков, для которых появление siRNA является сигналом к немедленному действию. На первом этапе с молекулой siRNA связываются белки-ферменты хеликаза и нуклеаза, формируя комплекс RISC (RNA-induced silencing complex; silence - англ. молчать, замолкать; silencing - замолкание, так в англоязычной и специальной литературе называют процесс «выключения» гена). Хеликаза раскручивает нити siRNA, в результате чего они расходятся. Одна из этих нитей, к которой прикреплен фермент нуклеаза, может теперь связаться с комплементарным участком однонитчатой мРНК, позволяя нуклеазе разрезать ее. Разрезанные же участки мРНК подвергаются действию других клеточных РНКаз, которые доразрезают их на более мелкие куски.
Итак, основная «специальность» siRNA в клетке - это блокирование тех генов, которые соответствуют одной из цепочек внутри siRNA. Но зачем механизм РНК - интерференции существует в клетках? Какие процессы он призван регулировать? Что собой представляют те мРНК, на которые направлено жало малых РНК, и почему их необходимо уничтожать?
Один из ответов на вопрос о необходимости РНК - интерференции в клетке, напрашивается сам собой: с помощью siRNA клетка может защищать себя от проникновения вирусов. Геном некоторых из этих опасных варваров состоит из ДНК, у некоторых же - из РНК, причем, против обычных правил, РНК у вирусов может быть как одно-, так и двунитчатой. Сам процесс разрезания чужеродной (вирусной) мРНК в этом случае происходит путем активаций комплекса ферментов RISC. Однако для большей эффективности растения и насекомые изобрели своеобразный путь усиления защитного действия siRNA. Присоединяясь к цепи мРНК, участок siRNA может с помощью комплекса ферментов, называемого DICER, сначала достроить вторую цепочку мРНК, а затем разрезать ее в разных местах, создавая, таким образом, разнообразные «вторичные» siRNA. Они, в свою очередь, формируют RISC и проводят мРНК через все стадии, о которых шла речь выше, вплоть до ее полного уничтожения. Такие «вторичные» молекулы смогут специфично связываться не только с тем участком вирусной мРНК, к которому была направлена «первичная» молекула, но также и с другими участками, что резко усиливает эффективность клеточной защиты.
Таким образом, у растений и низших животных организмов siRNA являются важным звеном своеобразного «внутриклеточного иммунитета», позволяющего распознавать и быстро уничтожать чужую РНК. В том случае, если в клетку проник РНК - содержащий вирус, такая система защиты не даст ему размножиться. Если же вирус содержит ДНК, система siRNA будет мешать ему производить вирусные белки (так как необходимая для этого мРНК будет распознаваться и разрезаться), и с помощью этой стратегии, замедлит его распространение по организму.
У млекопитающих же, в отличие от насекомых и растений, работает и другая система защиты. При попадании в «зрелую» (дифференцированную) клетку млекопитающего чужой РНК, длина которой больше 30 нуклеотидов, клетка начинает синтез интерферона. Интерферон, связываясь со специфическими рецепторами на клеточной поверхности, способен стимулировать в клетке целую группу генов. В результате в клетке синтезируется несколько видов ферментов, которые тормозят синтез белков и расщепляют вирусные РНК. Кроме того, интерферон может действовать и на соседние, еще не зараженные клетки, блокируя тем самым возможное распространение вируса.
Как можно заметить, обе системы во многом схожи: у них общая цель и «методы» работы. Даже сами названия «interferon» и «(RNA) interference» происходят от общего корня. Но есть у них и одно очень существенное различие: если интерферон при первых признаках вторжения просто «замораживает» работу клетки, не позволяя (на всякий случай) производство многих, в том числе и «невиновных» белков в клетке, то система siRNA отличается чрезвычайной разборчивостью: каждая siRNA будет распознавать и уничтожать только свою, специфическую мРНК. Замена всего лишь одного нуклеотида внутри siRNA ведет к резкому снижению эффекта интерференции.
В этом и заключается основное преимущество малых РНК: ни один из блокаторов генов, известных до сих пор, не обладает такой исключительной специфичностью по отношению к своему гену-мишени. Однако, как видно на примере многих опасных вирусных заболеваний у людей, ни иммунная, ни интерфероновая защита не всесильны, так что нам самое время позаимствовать у кого-нибудь передовой опыт в борьбе с вирусами. Почему бы не у растений или у насекомых? Ни те, ни другие не обладают системой адаптивного иммунитета. Чтобы выжить, растения были вынуждены «изобрести» РНК - интерференцию, которая до сих пор успешно защищает их клетки от внедрения вирусов. Появляется вполне закономерный вопрос: нельзя ли применить этот же подход в отношении клеток животных и людей?
Открытие siRNA, дает новую надежду в борьбе с вирусами, в том числе СПИДа, а также с теми, кого подозревают в соучастии при развитии рака. Высокая специфичность и эффективность вкупе с низкой токсичностью будут здесь как нельзя кстати. Как ожидается, применяя терапию siRNA вместе с традиционной антивирусной терапией, можно достичь эффекта потенцирования, когда два воздействия приводят к более выраженному лечебному эффекту, чем простая сумма каждого из них, применяемого по отдельности.
Для того, чтобы использовать механизм siRNA - интерференции в клетках млекопитающих, внутрь клеток нужно ввести уже готовые двухцепочечные молекулы siRNA. Оптимальный размер таких синтетических siRNA при этом составляет те же 21-28 нуклеотидов. Если увеличить ее длину - клетки ответят выработкой интерферона и снижением синтеза белка. Синтетические siRNA могут попасть как в зараженные, так и в здоровые клетки, и снижение выработки белков в незараженных клетках будет крайне нежелательным. С другой стороны, если попытаться применять siRNA меньшие, чем 21 нуклеотид, резко снижается специфичность ее связывания с нужной мРНК и способность к формированию комплекса RISC.
Так если удастся тем или иным способом доставить siRNA, обладающую способностью связываться с каким-либо участком генома ВИЧ (который, как известно, состоит из РНК), можно попытаться не допустить его встраивания в ДНК клетки хозяина. Кроме того, ученые разрабатывают пути воздействия на различные этапы размножения ВИЧ в уже зараженной клетке. Последний подход не обеспечит излечение, однако может существенно уменьшить скорость размножения вируса и дать загнанной в угол иммунной системе шанс «отдохнуть» от вирусной атаки, и самой попытаться расправиться с остатками заболевания.
Как предполагают, роль коротких РНК в клетке не ограничивается только лишь защитой от вирусов и других чужеродных РНК. Они, оказывается, были созданы, чтобы защитить нас... от нас самих.
Дело в том, что геном любого многоклеточного организма включает в себя множество элементов, которые когда-то были привнесены в него в процессе эволюции извне, например как результат встраивания вируса. Судите сами: из всего материала, содержащегося в наших с вами хромосомах, 34% приходится на долю элементов, называемых LINEs и SINEs (соответственно, Long и Short Interspersed Nuclear Elements), о функциях которых известно только то, что они могут по временам копировать себя и перемещаться с одного места хромосомы на другое; те участки ДНК, которые достались нам от ретровирусов (8% генома) и транспозоны (3%) также способны менять свое место в геноме. На их фоне всего лишь 2 (два(!)) процента собственно генов, кодирующих наши клеточные белки, кажутся такой же маловажной деталью, как и siRNA среди огромного разнообразия своих больших «сестер».
LINEs, SINEs, остатки вирусной ДНК и транспозоны, за свою способность к перемещениям именуемые подвижными, или мобильными элементами генома, представляют значительную опасность для наших хромосом. «Чужие среди своих», они при определенных обстоятельствах могут поднять бунт и привести к внутриклеточному хаосу. Некоторые из них - остатки вирусов, или протоонкогены - способны при «включении» вызывать рак; мобильные элементы, размножаясь и перемещаясь, меняют структуру хромосом, что может привести к мутациям. Например, у излюбленного объекта генетических исследований - плодовой мушки дрозофилы - более 80% спонтанных мутаций возникают именно из-за «хулиганского» поведения ее собственных мобильных элементов. Их перемещения внутри генома так индивидуальны и непредсказуемы, что положение некоторых из них может служить «молекулярным паспортом», точно определяющим личность хозяина, что уже используется на практике.
Понятно, что каждому организму необходимо было разработать систему жесткого контроля поведения мобильных элементов. Как раз эту «надзирательскую» функцию, как полагают ученые, и выполняет система внутриклеточных siRNA. На модели все того же С. elegans, например, было показано, что отключение генов, кодирующих некоторые из малых РНК, ведет к активизации перемещений мобильных элементов в его хромосомах и, соответственно, к повышению уровня мутаций.
Кроме того, ошибки в развитии органов и тканей при отключении генов, кодирующих систему siRNA у подопытных животных, а также ее активность в «незрелых» клетках указывают на то, что механизм РНК - интерференции активно участвует в регуляции программы «созревания» клеток и, как следствие, может играть одну из ключевых ролей в формировании целостного организма.
Еще одна из предполагаемых нормальных функций siRNA - отслеживание неправильно обработанных копий других типов РНК в клетке.
Наконец, в 2002 году исследователи неожиданно выяснили, что действие siRNA может не ограничиваться только лишь временным выключением генов на уровне РНК. Имеются данные, что в некоторых случаях siRNA, видимо, воздействует прямо на ДНК, изменяя структуру хроматина и способствуя длительному «замолканию» одних, и, возможно, активизации других генов.
Как осуществляются все эти функции siRNA, каковы их молекулярные детали? Откуда, например, может появляться та необходимая для запуска реакции РНК - интерференции молекула двухцепочечной РНК? Если она создается в клетке намеренно, то какие ферменты отвечают за ее создание? Какие свойства сигнализируют о том, что молекула одноцепочечной РНК «не своя» или ошибочная и подлежит уничтожению? Каковы особенности функционирования системы siRNA у разных организмов, и почему они возникли именно в таком виде? Как появляются siRNA в клетке млекопитающих, у которых пока не выявлены ферменты, ответственные за достраивание второй цепочки siRNA?
На эти и многие другие вопросы ответов пока нет. Каждый новый вопрос заставляет будущих профессоров и академиков, Нобелевских лауреатов и призеров Science искать ответ только лишь затем, чтобы задать новые вопросы следующим поколениям.
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 1268; Нарушение авторских прав?; Мы поможем в написании вашей работы!