КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Строение, функции и свойства ДНК. 3 страница
|
|
|
|
В ряде случаев в НК присутствуют редко встречающиеся минорные нуклеотиды, такие как дигидроуридин, 4-тиоуридин, инозин и др. Разнообразие их особенно велико у тРНК. Минорные нуклеотиды образуются в результате химических превращений оснований НК, происходящих уже после образования полимерной цепи. Чрезвычайно распространены в РНК и ДНК различные метилированные производные: 5-метилуридин, 5-метилцитидин, l-N-метиладенозин, 2-И-метилгуанозин. У РНК объектом метилирования могут быть и 2'-гидроксигруппы остатков рибозы, что приводит к образованию 2'-О-метилцитидина или 2'-О-метилгуанозина.
Рибонуклеотидные и дезоксирибонуклеотидные звенья соединяются между собой с помощью фосфодиэфирных мостиков, связывающих 5'-гидроксильную группу одного нуклеотида с 3'-гидроксильной группой следующего. Таким образом, регулярная основная цепь образована фосфатными и рибозными остатками, а основания присоединены к сахарам подобно тому, как присоединены боковые группы в белках. Порядок следования оснований вдоль цепи называется первичной структурой НК. Последовательность оснований принято читать в направлении от 5'- к 3'- углеродному атому пентозы.
Структура ДНК. Модель структуры ДНК в виде двойной спирали была предложена Уотсоном и Криком в 1953 г (рис.7).
Согласно этой трехмерной модели, молекула ДНК состоит из двух противоположно направленных полинуклеотидных цепей, которые относительно одной и той же оси образуют правую спираль. Азотистые основания находятся внутри двойной спирали, и их плоскости перпендикулярны основной оси, а сахарофосфатные остатки экспонированы наружу. Между основаниями образуются специфические Н-связи: аденин — тимин (или урацил), гуанин — цитозин, получившие название уотсон-криковского спаривания. В результате более объемные пурины всегда взаимодействуют с пиримидинами, имеющими меньшие размеры, что обеспечивает оптимальную геометрию остова. Антипараллельные цепи двойной спирали не являются идентичными ни по последовательности оснований, ни по нуклеотидному составу, но они комплементарны друг другу именно благодаря наличию специфического водородного связывания между указанными выше основаниями.
Комплементарность очень важна для копирования (репликации) ДНК. Соотношения между числом различных оснований в ДНК, выявленные

Рис.7. В - форма ДНК
Чарграффом с соавт. в 50-х гг., имели большое значение для установления структуры ДНК: было показано, что число адениновых остатков в основаниях цепи ДНК, независимо от организма, равно числу тиминовых, а число гуаниновых — числу цитозиновых. Эти равенства являются следствием избирательного спаривания оснований (рис.8).
Геометрия двойной спирали такова, что соседние пары оснований находятся друг от друга на расстоянии 0.34 нм и повернуты на 36° вокруг оси спирали. Следовательно, на один виток спирали приходится 10 пар оснований, и шаг спирали равен 3.4 нм. Диаметр двойной спирали равен 20 нм и в ней образуются два желобка — большой и малый. Это связано с тем, что сахарофосфатный остов расположен дальше от оси спирали, чем азотистые основания.
Стабильность структуры ДНК обусловлена разными типами взаимодействия, среди которых основными являются Н-связи между основаниями и межплоскостное взаимодействие (стэкинг). Благодаря последнему обеспечиваются не только выгодные ван-дер-ваальсовы контакты между атомами, но и возникает

Рис.8. Принцип комплементарности и антипараллельности цепей ДНК
дополнительная стабилизация вследствие перекрывания р-орбиталей атомов параллельно расположенных оснований. Стабилизации способствует также благоприятный гидрофобный эффект, проявляющийся в защищенности малополярных оснований от непосредственного контакта с водной средой. Напротив, сахарофосфатный остов с его полярными и ионизированными группами экспонирован, что также стабилизирует структуру.
Для ДНК известны четыре полиморфные формы: А, В, С и Z. Обычной структурой является В-ДНК, в которой плоскости пар оснований перпендикулярны оси двойной спирали (рис.7.). В А-ДНК плоскости пар оснований повернуты примерно на 20° от нормали к оси правой двойной спирали; на виток спирали здесь приходится 11 пар оснований. В С-ДНК на витке спирали 9 пар оснований. Z-ДНК — это левая спираль с 12 парами оснований на виток; плоскости оснований примерно перпендикулярны оси спирали. ДНК в клетке обычно находится в В-форме, но отдельные ее участки могут находиться в A, Z или даже в иной конформации.
Двойная спираль ДНК не застывшее образование, она находится в постоянном движении:
· деформируются связи в цепях;
· раскрываются и закрываются комплементарные пары оснований;
· ДНК взаимодействует с белками;
· если напряжение в молекуле велико, то она локально расплетается;
· правая спираль переходит в левую.
Различают 3 фракции ДНК:
1.Частоповторяемая (сателлитная) – до 106 копий генов (у мыши 10%). Она не участвует в синтезе белка; разделяет гены; обеспечивает кроссинговер; содержит транспозоны.
2.Слабоповторяемая – до 102 - 103 копий генов (у мыши 15%). Содержит гены синтеза т-РНК, гены синтеза белков рибосом и белков хроматина.
3.Уникальная (неповторяемая) – у мыши 75% (у человека 56%). Состоит из структурных генов.
Локализация ДНК: 95 % ДНК локализуется в ядре в хромосомах (линейные ДНК) и 5 % - в митохондриях, пластидах и клеточном центре в виде кольцевой ДНК.
Функции ДНК: хранение и передача информации; репарация; репликация.
Две цепи ДНК в области гена принципиально различаются по своей функциональной роли: одна из них является кодирующей, или смысловой, вторая — матричной.
Это значит, что в процессе «считывания» гена (транскрипции или синтеза пре-мРНК) в качестве матрицы выступает матричная цепь ДНК. Продукт же этого процесса-пре-мРНК — по последовательности нуклеотидов совпадает с кодирующей цепью ДНК (с заменой тиминовых оснований на урациловые).
Таким образом, получается, что с помощью матричной цепи ДНК при транскрипции воспроизводится в структуре РНК генетическая информация кодирующей цепи ДНК.
Главными матричными процессами, присущими всем живым организмам, являются репликация ДНК, транскрипция и трансляция.
Репликация — процесс, при котором информация, закодированная в последовательности оснований молекулы родительской ДНК, передается с максимальной точностью дочерней ДНК. При полуконсервативной репликации дочерние клетки первого поколения получают одну цепь ДНК от родителей, а вторая цепь является вновь синтезированной. Процесс осуществляется при участии ДНК-полимераз, которые относятся к классу трансфераз. Роль матрицы играют разделенные цепи двунитевой материнской ДНК, а субстратами являются дезоксирибонуклеозид-5'-трифосфаты.
Транскрипция — процесс переноса генетической информации от ДНК к РНК. Все виды РНК - мРНК, рРНК и тРНК - синтезируются в соответствии с последовательностью оснований в ДНК, служащей матрицей. Транскрибируется только одна, так называемая «+»-цепь ДНК. Процесс протекает при участии РНК-полимераз. Субстратами являются рибонуклеозид-5'-трифосфаты.
Процессы репликации и транскрипции у прокариот и эукариот существенно различаются по скорости протекания и по отдельным механизмам.
Трансляция — процесс декодирования мРНК, в результате которого информация с языка последовательности оснований мРНК переводится на язык аминокислотной последовательности белка. Осуществляется трансляция на рибосомах, субстратами являются аминоацил-тРНК.
Матричный синтез ДНК, катализируемый ДНК-полимеразами, выполняет две основные функции: репликацию ДНК — синтез новых дочерних цепей и репарацию двунитевых ДНК, имеющих разрывы в одной из цепей, образовавшихся в результате вырезания нуклеазами поврежденных участков этой цепи. У прокариот и эукариот существует три разновидности ДНК-полимераз. У прокариот выделены полимеразы I, II и III типов, обозначаемые как pol l, pol ll и pol III. Последняя катализирует синтез растущей цепи, pol играет важную роль в процессе созревания ДНК, функции pol ll изучены не полностью. В эукариотических клетках в репликации хромосом участвует ДНК-полимераза ά, в репарации — ДНК-полимераза β, а γ разновидность является ферментом, осуществляющим репликацию ДНК митохондрий. Эти Ферменты, независимо от типа клеток, в которых происходит репликация, присоединяют нуклеотид к ОН-группе на З'-конце одной из цепей ДНК, которая растет в направлении 5'→3. Поэтому говорят, что данные Ф обладают 5'→3'-полимеразной активностью. Помимо этого все они проявляют способность деградировать ДНК, отщепляя, нуклеотиды в направлении 3'→5, т. е. являются 3'→5'-экзонуклеазами.
В 1957 г. Мезельсон и Сталь, изучая E. coli установили, что на каждой свободной цепи фермент ДНК-полимераза строит новую, комплементарную цепь. Это полуконсервативный способ репликации: одна цепь старая – другая новая!
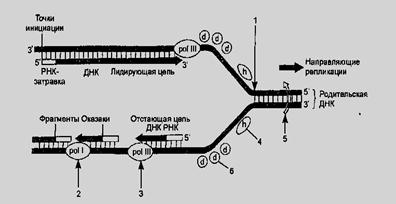
Обычно репликация начинается в строго определенных участках, получивших название участков ori (от origin of replication), и от этих участков распространяется в обе стороны. Участкам ori предшествуют точки разветвления материнских цепей ДНК. Участок, примыкающий к точке разветвления, получил название репликативной вилки (рис.9). В ходе синтеза репликативная вилка перемещается вдоль молекулы, при этом расплетаются все новые участки родительской ДНК до тех пор, пока вилка не дойдет до точки терминации. Разделение цепей достигается с помощью специальных Ф — геликаз (топоизомераз). Энергия, необходимая для этого, высвобождается за счет гидролиза АТФ. Геликазы перемещаются вдоль полинуклеотидных цепей в двух направлениях.
Для начала синтеза ДНК необходима затравка — праймер. Роль праймера выполняет короткая РНК (10—60 нуклеотидов). Она синтезируется комплементарно определенному участку ДНК при участии праймазы. После образования праймера в работу включается ДНК-полимераза. В отличие от геликаз ДНК-полимеразы могут перемещаться только от 3' к 5' концу матрицы. Поэтому элонгация растущей цепи по мере раскручивания двунитевой материнской ДНК может идти только вдоль одной цепи матрицы, той, относительно которой вилка репликации движется от 3' к 5' концу. Непрерывно синтезируемая цепь получила название лидирующей. Синтез на запаздывающей цепи также начинается с образования праймера и идет в направлении, противоположном ведущей цепи — от вилки репликации. Запаздывающая цепь синтезируется фрагментарно (в виде фрагментов Оказаки), т. к. праймер образуется только тогда, когда вилка репликации освободит тот участок матрицы, который имеет сродство к праймазе. Лигирование (сшивание) фрагментов Оказаки с образованием единой цепи носит название процесса созревания.
При созревании цепи РНК-затравка удаляется как с 5' конца ведущей цепи, так и с 5' концов фрагментов Оказаки, а эти фрагменты сшиваются друг с другом. Удаление затравки осуществляется при участии 3'→5' экзонуклеазы. Этот же Ф вместо удаленной РНК присоединяет дезоксинуклеотиды, используя свою 5'→3' полимеразную активность. При этом в случае присоединения «неправильного» нуклеотида осуществляется «корректорская правка» — удаление оснований, образующих некомплементарные пары. Этот процесс обеспечивает чрезвычайно высокую точность репликации, отвечающую одной ошибке на 109 пар оснований.
Рис.9. Репликация ДНК:
1 — репликативная вилка, 2 — ДНК-полимераза (pol I — созревание);
3 — ДНК-полимераза (pol III — «корректорская правка»); 4—геликаза;
5—гираза (топоизомераза); 6—белки, дестабилизирующие двойную спираль.
 |
Коррекция осуществляется в тех случаях, когда к З'-концу растущей цепи присоединяется «неправильный» нуклеотид, неспособный образовать нужные водородные связи с матрицей. Когда pol III ошибочно присоединяет неправильное основание, «включается» ее 3' -» 5'-экзонуклеазная активность, и это основание немедленно удаляется, после чего восстанавливается полимеразная активность. Такой простой механизм действует благодаря тому, что pol III способна работать как полимераза лишь на совершенной двойной спирали ДНК с абсолютно правильным спариванием оснований.
Еще один механизм удаления РНК-фрагментов основан на присутствии в клетках особой рибонуклеазы, получившей название РНКазы Н. Этот Ф специфичен к двунитевым структурам, построенным из одной рибонуклеотидной и одной дезоксирибонуклеотидной цепи, причем он гидролизует первую из них.

 РНКаза Н также способна удалять РНК-праймер с последующей застройкой разрыва с помощью ДНК-полимеразы. На заключительных этапах сборки фрагментов в нужном порядке действует ДНК-лигаза, катализирующая образование фосфодиэфирной связи.
РНКаза Н также способна удалять РНК-праймер с последующей застройкой разрыва с помощью ДНК-полимеразы. На заключительных этапах сборки фрагментов в нужном порядке действует ДНК-лигаза, катализирующая образование фосфодиэфирной связи.
Раскручивание геликазами части двойной спирали ДНК в хромосомах эукариот приводит к сверхспирализации остальной части структуры, что неизбежно сказывается на скорости процесса репликации. Сверхспирализации препятствуют ДНК-топоизомеразы.
Таким образом, в репликации ДНК, помимо ДНК-полимеразы, принимает участие большой набор Ф: геликаза, праймаза, РНКаза Н, ДНК-лигаза и топоизомераза. Этим перечень Ф и белков, участвующих в матричном биосинтезе ДНК, далеко не исчерпывается. Однако многие из участников этого процесса до настоящего времени остаются мало изученными.
 В процессе репликации происходит «корректорская правка» - удаление неправильных (образующих некомплементарные пары) оснований, включенных во вновь синтезированную ДНК. Этот процесс обеспечивает чрезвычайно высокую точность репликации, отвечающую одной ошибке на 109 пар оснований.
В процессе репликации происходит «корректорская правка» - удаление неправильных (образующих некомплементарные пары) оснований, включенных во вновь синтезированную ДНК. Этот процесс обеспечивает чрезвычайно высокую точность репликации, отвечающую одной ошибке на 109 пар оснований.
Теломеры. В 1938г. классики генетики Б.Мак-Клинтон и Г. Мёллер доказали, что на концах хромосом есть специальные структуры, которые назвали теломерами (телос-конец, мерос-часть).
Ученые обнаружили, что при воздействии рентгеновским облучением устойчивость проявляют лишь теломеры. Напротив, лишенные концевых участков, хромосомы начинают сливаться, что ведет к тяжелым генетическим аномалиям. Т.о., теломеры обеспечивают индивидуальность хромосом. Теломеры плотно упакованы (гетерохроматин) и малодоступны для ферментов (теломеразы, метилазы, эндонуклеаз и др.)
Функции теломер.
1.Механические: а) соединение концов сестринских хроматид после S-фазы; б) фиксация хромосом к ядерной мембране, что обеспечивает конъюгацию гомологов.
2.Стабилизационные: а) предохранение от недорепликации генетически значимых отделов ДНК (теломеры не транскрибируются); б) стабилизация концов разорванных хромосом. У больных α - талассемией в генах α - глобина происходят разрывы хромосомы 16д и к поврежденному концу добавляются теломерные повторы (ТТАГГГ).
3.Влияние на экспрессию генов. Активность генов, расположенных рядом с теломерами, снижена. Это проявление сайленсинга – транскрипционное молчание.
4.«Счетная функция». Теломеры выступают в качестве часового устройства, которое отсчитывает количество делений клетки. Каждое деление укорачивает теломеры на 50-65 н.п. А всего их длина в клетках эмбриона человека составляет 10-15 тысяч н.п.
Теломерная ДНК попала в поле зрения биологов совсем недавно. Первые объекты исследования – одноклеточные простейшие – ресничная инфузория (тетрахимена), которая содержит несколько десятков тысяч очень мелких хромосом и, значит, множество теломер в одной клетке (у высших эукариот менее 100 теломер на клетку).
В теломерной ДНК инфузории многократно повторяются блоки из 6-ти нуклеотидных остатков. Одна цепь ДНК содержит блок 2 тимин – 4 гуанин (ТТГГГГ - Г-цепь), а комплементарная цепь - 2 аденин – 4 цитозин (ААЦЦЦЦ - Ц-цепь).
Каково же было удивление ученых, когда обнаружили, что теломерная ДНК человека отличается от таковой у инфузории всего лишь одной буквой и образует блоки 2 тимин – аденин – 3 гуанин (ТТАГГГ). Более того, оказалось, что из ТТАГГГ - блоков построены теломеры (Г – цепь) всех млекопитающих, рептилий, амфибий, птиц и рыб.
Впрочем, удивляться здесь нечему, так как в теломерной ДНК не закодировано никаких белков (она не содержит гены). У всех организмов теломеры выполняют универсальные функции, речь о которых шла выше. Очень важная характеристика теломерных ДНК – их длина. У человека она колеблется от 2 до 20 тысяч пар оснований, а у некоторых видов мышей может достигать сотен тысяч н.п. Известно, что около теломер есть специальные белки, обеспечивающие их работу и участвующие в построении теломер.
Доказано, что для нормального функционирования каждая линейная ДНК должна иметь две теломеры: по одной теломере на каждый конец.
У прокариот теломеров нет – их ДНК замкнута в кольцо.
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 2303; Нарушение авторских прав?; Мы поможем в написании вашей работы!