КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Строение, функции и свойства ДНК. 2 страница
|
|
|
|
Биорегуляторы - естественные регуляторы жизненно важных функций человеческого организма. Эта обширная группа биологически активных веществ, которая контролирует сердечную деятельность, дыхание, тонус мышц и сосудов, пищеварение, иммунитет, а также высшую психическую деятельность - эмоции, мышление, память. Биорегуляторы еще часто называют «гормональным» или «биохимическим» оружием.
Четкого определения термина «биорегулятор», как и классификации, на сегодняшний день не существует. Слишком многочисленна и разнообразна эта группа веществ. К биорегуляторам относят гормоны, цитокинины, простагландины, лейкотриены, медиаторы, ферменты и многие другие вещества. Наиболее общим признаком для всех этих соединений является высокая биологическая активность в сверхмалых дозах -1x10-15 и даже 1x10-18 молей.
Рецепторная функция. Белковую природу имеют мембранные рецепторы для физиологически активных соединений, которые принимают химический сигнал от гормонов, нейромедиаторов (адренорецепторы, холинорецепторы, гистаминовые рецепторы и др.), биологически активных веществ (БАВ) и др. Для врача и фармацевта очень важно знать принцип взаимодействия ЛЕКАРСТВО-РЕЦЕПТОР.
Клетки, как правило, содержат большое количество различных мембранных, цитоплазматических и ядерных рецепторов БАВ: ионные каналы, ферменты, нейромедиаторы, транспортные системы и гены. Они необходимы организму для нормального функционирования и адекватного реагирования на изменения внешней и внутренней среды и, в конечном счете, для поддержания гомеостаза. Наличие у живых существ органов, воспринимающих внешние сигналы, было отмечено учеными еще в глубокой древности. Однако понимание роли рецепторов как внутренней системы «органов чувств», позволяющей организмам адаптироваться к окружающей среде, стало складываться только в середине XIX в. Основоположником рецепторной теории действия лекарств следует признать P. Ehrlich (1907 г.). Он же впервые ввел термин «рецептор» и сформулировал основной постулат: «corpora non agun nisi fixata» — «вещества не действуют, если не фиксируются». Это обобщение стало теоретической базой химиотерапии. P. Ehrlich предполагал, что клетки содержат так называемые боковые ветви, или рецепторы, в результате селективного взаимодействия с которыми ЛВ индуцируют свой эффект. Согласно этой теории, лекарство имеет два структурных фрагмента, один из которых, соединяясь с рецептором, позволяет тем самым другому фрагменту осуществлять биологический ответ. Многочисленные исследования подтвердили правильность теории P. Ehrlich о рецепторах как участках тканей, селективно связывающих ЛВ и опосредующих реализацию их фармакологических эффектов.
В обобщенном виде понятие «рецептор» можно сформулировать следующим образом: рецептором может быть любая высокомолекулярная конформационно подвижная биоструктура, специфически связывающая химическое соединение (лиганд, лекарство) на поверхности или внутри клетки и трансформирующая полученную информацию в биологический ответ. К таким макромолекулам относятся белки и нуклеиновые кислоты. Специфическое взаимодействие лиганда с рецептором следует отличать от неспецифического связывания эндогенных или экзогенных БАВ с белками плазмы крови или мукополисахаридами соединительной ткани. Белковые структуры такого типа получили название «молчащих» рецепторов. В результате связывания с ними не реализуются никакие эффекты. Плазменные белки количественно, а не качественно влияют на проявление эффектов ЛВ.
К началу 90-х гг. было выявлено более 30 групп различных рецепторов. В последнее десятилетие, благодаря расшифровке генома человека, этот список значительно расширился и продолжает пополняться.
Во всем многообразии рецепторов выделяют 4 следующих типа:
· рецепторы, осуществляющие контроль за функцией ионных каналов (н-холинорецепторы, GАВАА-рецепторы, глутаматные рецепторы);
· рецепторы, сопряженные с эффектором через систему G-протеины — вторичные посредники или G-протеины — ионные каналы (рецепторы некоторых белковых гормонов и пептидов — ангиотензина, брадикинина, эндотелина и др.; биогенных аминов — адреналина, дофамина, гистамина, серотонина; липидов — канабиноидов, простагландинов, лейкотриенов, тромбоксанов; нуклеозидов и нуклеотидов — аденозина, АТФ, АДФ и др., а также ионов Са2+);
· рецепторы, осуществляющие прямой контроль за функцией эффекторного фермента (они непосредственно связаны с тирозинкиназой и регулируют фосфорилирование белков — рецепторы инсулина и гормонов роста);
· рецепторы, контролирующие транскрипцию ДНК — растворимые цитозольные или ядерные белки (рецепторы стероидных и тиреоидных гормонов).
Первые три типа рецепторов принадлежат к числу мембранных, а последний — к ядерным. Большинство рецепторов представляют собой белки, точнее липо- или гликопротеины, которые располагаются на цитоплазматической мембране или мембранах органелл клетки. Они могут пронизывать всю толщу мембраны или находиться с ее внутренней стороны. Если лиганды рецептора представляют собой гидрофильные молекулы, например, нейромедиаторы, то связывающие участки рецептора должны обязательно находиться с внешней стороны клетки. Рецепторы гидрофобных молекул слабо связаны с мембранами, и образующиеся комплексы с лигандами могут выходить из мембраны в цитоплазму.
В процессе эволюции образовались также рецепторы, локализованные внутри клеток. Их лиганды, например, стероидные или тиреоидные гормоны, обладают способностью за счет своих гидрофобных свойств проникать через мембраны путем пассивной диффузии или активного транспорта. Структура и функции внутренних рецепторов менее изучены, чем мембранных, поскольку первые труднее выделить, не изменив при этом их первоначальной структуры. Известно присутствие рецепторов различных лигандов па рибосомах (к антибиотикам), в ядре (к стероидным и тиреоидным гормонам, тиазолидиндионам, жирным эссенциальным кислотам, фибратам), в комплексе Гольджи (к гормонам), в микросомах (к гормонам). Биологический смысл такой внутриклеточной рецепции заключается в более глубоком и продолжительном изменении функций клетки в ответ на получение внешнего сигнала.
Белковые цепи и домены, образующие рецептор, имеют определенную организацию. Установлено, что в механизме, обеспечивающем закрытие ионного канала, ведущая роль принадлежит н-ацетилхолинорецептору, состоящему из пяти белковых субъединиц, каждая из которых имеет четыре трансмембранных домена. Последние образованы пронизывающими всю мембрану гидрофобными a-спиралями. Один из доменов — М2, выстилающий канал с внутренней стороны, гидрофилен. В нем преобладают ориентированные особым образом анионсодержащие фрагменты аминокислот, что обеспечивает распознавание катионов. При взаимодействии лиганда с таким рецептором, за счет изменений в трансмембранном домене М2, происходит конформационная перестройка белков, образующих стенки канала, и закрытие его просвета.
Представление о строении рецепторов, передача сигналов от котoрых осуществляется с участием G-протеинов, можно составить по (рис.3) из которого видно, что единственная полипептидная цепь имеет семь трансмембранных а-спиралей, а находящаяся вне мембраны цепь с терминальной NН2-группой, гликозилирована. Петли, выходящие в цитоплазму клетки, различаются по количеству аминокислотных остатков. Первая состоит из 10—20 аминокислот, вторая содержит до 20 и третья — до 50 аминокислотных остатков. На третьей петле расположены участки связывания с G-протеинами. Оканчивается G-сопряженный рецептор цепью из 100 аминокислот и содержит концевую карбоксильную группу, которая также обеспечивает связывание с G-белками. Сведения о пространственном строении рецепторов этого типа стали доступны лишь недавно. Благодаря рентгено-структурному исследованию (РСИ) бычьего родопсина была впервые предложена трехмерная (3D) модель рецептора, сопряженного с G-белками.
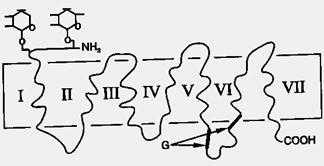
Рис.3. Схематическое изображение рецептора, связанного с G-белками: G — домены, обеспечивающие связывание с G-протеинами
В настоящее время в результате расшифровки генома человека выявлено несколько сотен рецепторов, сопряженных с G-белками. Из них только 30 являются мишенями широко распространенных ЛВ, а для 210 — идентифицированы лишь эндогенные лиганды. К этим группам принадлежат рецепторы пептидов, биогенных аминов, нуклеозидов и лейкотриенов. Примерно 160 рецепторов, выявленных в человеческом геноме, относят к так называемым «рецепторам-сиротам», т. к. их лиганды и физиологические функции пока не установлены.
Обособленную группу образует семейство рецепторов, в передаче сигналов от которых участвует тирозинкиназа. К их числу принадлежат инсулиновые рецепторы и рецепторы факторов роста. Тирозиновый домен находится внутри клетки. Экстрацеллюлярная часть содержит области, богатые цистеиновыми остатками, которые, вероятно, и обеспечивают вторичную структуру, необходимую для мест связывания лигандов.
Характерной особенностью внутриклеточных рецепторов стероидных гормонов является наличие домена, способного связывать полипептидные цепи с лигандом за счет Zn-пальцев: цистеиновые остатки окружают атомы Zn и обеспечивают связь с ДНК.
Взаимодействие лигандов с рецепторами осуществляется благодаря образованию либо водородных и координационных связей, либо комплексов с переносом заряда, ион-ионного, ион-дипольного или дисперсионного взаимодействия.
Транспортная функция. Белки связывают и осуществляют межклеточный и внутриклеточный (трансмембранный цитоплазматический) транспорт разных лигандов – биомолекул, ионов металлов, чужеродных химических соединений (ксенобиотиков). Транспортными белками крови являются сывороточные альбумины (переносят жирные кислоты, билирубин, лекарственные и токсические соединения), гемоглобин эритроцитов (транспортирует кислород), липопротеины (транспортируют липиды), трансферин (транспортирует железо).

Рис.4. Гемоглобин (тетрамер) с гемом в центре
Гемоглобин- это белок, переносящий кислород от легких к тканям и осуществляющий транспорт углекислого газа от тканей обратно к легким. Гемоглобин локализован в красных кровяных клетках-эритроцитах. Молекула гемоглобина состоит из четырех полипептидных цепей- двух идентичных α-цепей (обозначаемых α1 и α2) и двух идентичных β-цепей (обозначаемых β1 и β2). Каждая цепь связана с особой группой- гемом (рис.4).
Миоглобин- это белок, переносящий кислород в мышечных клетках. Он состоит из одной полипептидной цепи и имеет один гем.
Третичная структура α- и β-цепей гемоглобина и цепи миоглобина оказалась сходной. Гем состоит из атомов углерода, азота и водорода, образующих плоское кольцо, называемое порфирином. В центре этого кольца находится атом железа, связанный с атомами кольца четырьмя из шести своих возможных координационных связей.
Дезоксигемоглобин- это гемоглобин, не связанный с кислородом. В дезоксигемоглобине атом железа находится вне плоскости порфиринового кольца и, кроме того, имеется несколько солевых мостиков, и водородных связей, которых нет в оксигенированной форме белка.
Оксигемоглобин-это гемоглобин, связанный с кислородом. Кислород занимает шестое координационное положение атома железа гема и располагается рядом с дальним остатком гистидина. При связывании кислорода атом железа перемещается в плоскость порфиринового кольца. Одновременно с этим происходят и некоторые другие конформационные изменения.
Гемоглобин плода человека (фетальный гемоглобин) отличается от гемоглобина взрослых тем, что в нем две β-цепи заменены двумя γ-цепями. Гемоглобин плода связывает ДФГ менее сильно, чем дезоксигемоглобин взрослых, и поэтому имеет более высокое сродство к кислороду. В результате, когда кровь плода обогащается кислородом за счет материнской крови, что имеет место в плаценте, кислород переходит от матери к плоду с высокой эффективностью.
Серповидноклеточная анемия- заболевание, возникающее при замене остатка глутаминовой кислоты в шестом положении β-цепи гемоглобина на остаток валина. Это единственное изменение приводит к уменьшению растворимости дезоксигемоглобина и к появлению у него способности агрегировать с образованием длинных волокон. В результате эритроциты приобретают вытянутую форму, напоминающую серп. Деформированные таким образом, они могут закупоривать капилляры, нарушая нормальное кровообращение, либо легко подвергаться лизису, что и является непосредственной причиной наблюдаемой анемии.
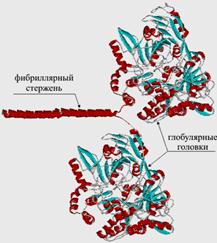 |
Рис.5. Схематическое строение белка миозина
Сократительная функция. Белки являются молекулярными структурами, которые реализуют сократительную функция мышц (актин, миозин), жгутиков и ресничек (тубулины, динеины). Сократительные и двигательные белки придают организму способность сокращаться, изменять форму и перемещаться, прежде всего, речь идет о мышцах. 40% от массы всех белков, содержащихся в мышцах, составляет миозин (mys, myos, греч. – мышца). Его молекула содержит одновременно фибриллярную и глобулярную часть (рис.5).
Такие молекулы объединяются в крупные агрегаты, содержащие 300–400 молекул. При изменении концентрации ионов кальция в пространстве, окружающем мышечные волокна, происходит обратимое изменение конформации молекул – изменение формы цепи за счет поворота отдельных фрагментов вокруг валентных связей. Это приводит к сокращению и расслаблению мышц, сигнал для изменения концентрации ионов кальция поступает от нервных окончаний в мышечных волокнах. Искусственное сокращение мышц можно вызвать действием электрических импульсов, приводящих к резкому изменению концентрации ионов кальция, на этом основана стимуляция сердечной мышцы для восстановления работы сердца.
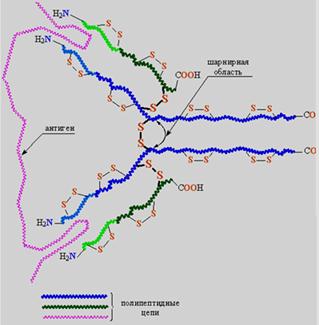
Рис.6. Схематическое изображение структуры иммуноглобулина
Защитная функция. Белки выполняют функцию иммунной защиты (иммуноглобулины, лимфокины, интерлейкины), противодействуют кровотечению и тромбообразованию (белки сворачивающейся, антикоагулянтной и фибринолитической систем крови).
Защитные белки позволяют уберечь организм от вторжения атакующих его бактерий, вирусов и от проникновения чужеродных белков (обобщенное название чужеродных тел – антигены). Роль защитных белков выполняют иммуноглобулины (другое их название – антитела), они распознают антигены, проникшие в организм, и прочно связываются с ними. В организме млекопитающих, включая человека, есть пять классов иммуноглобулинов: M, G, A, D и E.
Их структура, как следует из названия, глобулярная, кроме того, все они построены сходным образом. Молекулярная организация антител показана на примере иммуноглобулина класса G (рис.6). Молекула содержит четыре полипептидные цепи, объединенные тремя дисульфидными мостиками S-S (на рис. они показаны с утолщенными валентными связями и крупными символами S), кроме того, каждая полимерная цепь содержит внутрицепные дисульфидные перемычки. Две большие полимерные цепи содержат 400–600 аминокислотных остатков. Две другие цепи почти вдвое короче, они содержат приблизительно 220 аминокислотных остатков. Все четыре цепи расположены таким образом, что концевые H2N-группы направлены в одну сторону.
Молекула состоит из четырех полимерных цепей, соединенных дисульфидными мостиками -S-S-, которые отмечены утолщенными валентными связями и крупными символами S. Области, захватывающие инородный белок – антиген, выделены светло-синим и светло-зеленым цветом. В центральной части молекулы находится шарнирная область, которая обеспечивает независимую подвижность областей захвата. Фрагмент реальной структуры иммуноглобулина показан в нижней части рисунка, справа. Это глобулярная молекула, содержащая b-структуры, соединенные одиночными цепями.
После контакта организма с чужеродным белком (антигеном), клетки иммунной системы начинают вырабатывать иммуноглобулины (антитела), которые накапливаются в сыворотке крови. На первом этапе основную работу совершают участки цепей, содержащие концевые H2N. Это области захвата антигенов. В процессе синтеза иммуноглобулина эти участки формируется таким образом, чтобы их строение и конфигурация максимально соответствовали структуре приблизившегося антигена (как ключ к замку, подобно ферментам, но задачи в данном случае иные). Таким образом, для каждого антигена в качестве иммунного ответа создается строго индивидуальное антитело. Столь «пластично» изменять строение в зависимости от внешних факторов, помимо иммуноглобулинов, не может ни один известный белок. Ферменты решают задачу структурного соответствия реагенту иным путем – с помощью гигантского набора разнообразных ферментов в расчете на все возможные случаи, а иммуноглобулины каждый раз заново перестраивают «рабочий инструмент». Сверх того, шарнирный участок иммуноглобулина обеспечивает двум областям захвата некоторую независимую подвижность, в результате молекула иммуноглобулина может «найти» сразу два наиболее удобных для захвата участка в антигене с тем, чтобы его надежно зафиксировать.
Далее включается цепь последовательных реакций иммунной системы организма, подключаются иммуноглобулины других классов, в результате происходит дезактивация чужеродного белка, а затем уничтожение и удаление антигена (постороннего микроорганизма или токсина).
После контакта с антигеном максимальная концентрация иммуноглобулина достигается (в зависимости от природы антигена и индивидуальных особенностей самого организма) в течение нескольких часов (иногда нескольких дней). Организм сохраняет память о таком контакте, и при повторной атаке таким же антигеном иммуноглобулины накапливаются в сыворотке крови значительно быстрее и в большем количестве – возникает приобретенный иммунитет.
Иммуноглобулин G. Приблизительно 75% иммуноглобулинов плазмы представлены IgG — основным классом антител, продуцируемых при вторичном ответе. IgG лучше, чем другие иммуноглобулины, диффундирует во внесосудистое пространство, а также проходит через плаценту, обеспечивая пассивный иммунитет новорожденным в первые недели жизни. Иммуноглобулины нейтрализуют бактериальные токсины и связываются с микроорганизмами, усиливают фагоцитоз.
Иммуноглобулин A — основной класс антител в секретах, таких, как слеза, слюна, респираторные секреты, мочеполовые и желудочно-кишечные секреты. Иммуноглобулины существуют в виде одиночных иммуноглобулиновых единиц или в виде димеров, связанных секреторной единицей (ММ 60 000) и дополнительной пептидной цепью (J-цепь, ММ 20000). IgA покрывают микроорганизмы и угнетают их способность прикрепляться к поверхности слизистых оболочек.
Иммуноглобулин M — основной класс антител, секретируемых на ранних стадиях первичного иммунного ответа. Эти иммуноглобулины являются полимерами пяти иммуноглобулиновых единиц, объединенных вокруг J-цепи. Они в основном прикреплены к поверхности сосудов и являются эффективными цитолитическими антителами. После рождения уровни IgM прогрессивно растут, так как новорожденные подвергаются антигенному стимулированию. Определение IgM у новорожденных используется как индикатор внутриматочной инфекции.
Иммуноглобулин D определен на поверхности неактивированных В-лимфоцитов, где он, вероятно, выполняет функцию рецепторов для антигенов. Концентрация иммуноглобулинов данного типа в плазме очень низка.
Иммуноглобулин Е представлен антителами, связанными с поверхностью тучных клеток. В плазме IgE присутствуют в незначительных количествах. Связывание антигена приводит к дегрануляции тучных клеток с высвобождением активных аминов (гистамин), повышающих реакции гиперчувствительности.
Приведенная функциональная классификация белков носит в определенной степени условный характер, например белок тромбин, упомянутый среди защитных белков, по существу представляет собой фермент, катализирующий гидролиз пептидных связей, то есть, относится к классу протеаз.
Белки как источники питания. Белки в живом организме постоянно расщепляются на исходные аминокислоты (с непременным участием ферментов), одни аминокислоты переходят в другие, затем белки вновь синтезируются (также с участием ферментов), т.е. организм постоянно обновляется. Некоторые белки (коллаген кожи, волос) не обновляются, организм непрерывно их теряет и взамен синтезирует новые. Белки как источники питания выполняют две основные функции: они поставляют в организм строительный материал для синтеза новых белковых молекул и, кроме того, снабжают организм энергией (источники калорий). Плотоядные млекопитающие (в том числе и человек) получают необходимые белки с растительной и животной пищей. Ни один из полученных с пищей белков не встраивается в организм в неизменном виде. В пищеварительном тракте все поглощенные белки расщепляются до аминокислот, и уже из них строятся белки, необходимые конкретному организму, при этом из 8 незаменимых кислот в организме могут синтезироваться остальные 12, если они не поступают в достаточном количестве с пищей, но незаменимые кислоты должны поступать с пищей непременно. Атомы серы в цистеине организм получает с незаменимой аминокислотой – метионином. Часть белков распадается, выделяя энергию, необходимую для поддержания жизнедеятельности, а содержавшийся в них азот выводится из организма с мочой. Обычно организм человека теряет 25–30 г. белка в сутки, поэтому белковая пища должны постоянно присутствовать в нужном количестве. Минимальная суточная потребность в белке составляет у мужчин 37 г, у женщин 29 г, однако рекомендованные нормы потребления почти вдвое выше. При оценке пищевых продуктов важно учитывать качество белка. При отсутствии или низком содержании незаменимых аминокислот белок считается малоценным, поэтому такие белки должны потребляться в большем количестве. Так, белки бобовых культур содержат мало метионина, а в белках пшеницы и кукурузы низкое содержанием лизина (обе аминокислоты незаменимые). Животные белки (исключая коллагены) относят к полноценным пищевым продуктам. Полный набор всех незаменимых кислот содержит казеин молока, а также приготовляемые из него творог и сыр, поэтому вегетарианская диета, в том случае, если она очень строгая, т.е. «безмолочная», требует усиленного потребления бобовых культур, орехов и грибов для снабжения организма незаменимыми аминокислотами в нужном количестве.
Конец XX и начало XXI столетий ознаменовались величайшими открытиями в области молекулярной биологии и генетики, которые революционым образом изменили наше представление о механизмах кодирования и реализации наследственной информации у эукариот, создали возможности для изучения генов и геномов.
В настоящее время можно говорит о создании нового направления в молекулярной генетике - геномики. К числу открытий геномики относятся расшифровка геномов многих микроорганизмов, ряда многоклеточных, в том числе лабораторных животных (дрозофила, мышь), и, наконец, практически полная расшифровка генома человека. Оказалось, что этот геном содержит значительно меньшее количество генов, чем предполагалось ранее. По предварительным оценкам, в геноме человека от 22000 до 25000 структурных генов. Однако они имеют чрезвычайно сложное строение и механизмы экспрессии. Классическая гипотеза «один ген - один фермент» претерпела существенные изменения: найдены механизмы кодирования нескольких белков одним геном. Общее количество синтезируемых у человека белков в периоде онтогенеза предположительно достигает 250 000. Совокупность белков, синтезируемых организмом в течение жизни, получила название протеом. В настоящее время понятно, что только изучение протеома позволит уточнить функции многих генов и завершит изучение генома человека. Поэтому появилось второе важное направление современной молекулярной биологии - протеомика. Слово «протеом» образовано от слова «протеин» (белок) и окончания слова «геном», так что в самом названии как бы слиты воедино белок и геном (ДНК). Это подчеркивает их теснейшую взаимосвязь. Однако между геномикой и протеомикой, между геномом и протеомом есть одно фундаментальное различие, которое вызывает к жизни совершенно новые методы исследования, новые стратегии.
Протеом - понятие динамическое, тогда как геном стабилен и постоянен, иначе было бы невозможно передать наследственные свойства от поколения к поколению, обеспечить сохранение видов и т.д. Изменчивость генома всегда происходит на фоне его высокой стабильности и ни в коей мере ее не отменяет. Протеом - набор белков данной клетки в данной фазе ее развития в данный момент времени - меньше генома по общему объему информации. В любой клетке человеческого организма никогда не функционируют все 22-25 тыс. генов, работает лишь их часть - иногда меньшая, иногда большая. Хотя точные цифры привести пока трудно, но в обычной специализированной клетке, например клетке печени или легкого, одновременно присутствуют, вероятно, не более 10 тыс. белков, причем в резко различных количествах - от нескольких молекул на клетку до нескольких процентов общего клеточного белка. Набор белков постоянно меняется в зависимости от фазы клеточного деления, тканевой специализации клетки, стадии ее дифференцировки, принадлежности к нормальным или злокачественным клеткам, состояния стресса или покоя, воздействия внеклеточных физиологически активных веществ и так до бесконечности. Поэтому белковый «портрет» клетки зависит от множества факторов и воздействий, подвержен практически непрерывным изменениям, что делает его изучение особенно трудным.
Изменчивость протеома связана не только с тем, что в данный момент времени работает одна часть генов, а в другой момент - иная. Набор белков сильно зависит от процессов, протекающих на пути от ДНК к матричной РНК (мРНК). Здесь большая часть первичных генных продуктов (РНК) подвергается так называемому альтернативному сплайсингу, суть которого состоит в том, что до образования зрелой матричной РНК из нее удаляются разные части молекулы. В результате один ген может породить множество белков, различающихся первичной структурой. Таким образом, стало очевидно, что одна из старых догм биохимии и молекулярной биологии - «Один ген - один фермент» - нуждается в модернизации. Для очень многих случаев справедлива формула: «Один ген - много белков».
Однако и это еще не все. После синтеза белки претерпевают множество химических изменений (модификаций), которые создают их огромное разнообразие, хотя исходно они кодированы одним геном. К числу таких модификаций относятся реакции фосфорилирования, ацетилирования, метилирования, гликозилирования и многие другие. Если учесть, что на большом белке есть множество мест, где эти модификации могут происходить, то легко себе представить, какое практически бесконечное разнообразие форм одной и той же белковой молекулы может возникнуть. Подавляющее большинство модификаций существенно сказывается на биологической активности данной молекулы белка, а также на ее способности взаимодействовать с другими белковыми молекулами. В итоге мы приходим к заключению, что когда в клетке работает, скажем, 10% всех генов, допустим 8 тыс., то количество разных белков может превысить эту величину в 10 раз. Исследователи и раньше догадывались, что такая ситуация возможна, однако только теперь реально представляют ее истинные масштабы. Крайне важным разделом протеомики, безусловно, следует считать изучение белок - белковых и белок - нуклеиновых взаимодействий. В течение жизни клетки практически каждый белок при своем функционировании взаимодействует с множеством макромолекул, а также низкомолекулярных лигандов. В связи с этим, один из руководителей проекта «Геном человека» Дж. Крейг Вентер считает, что для полного определения функций всех генов может потребоваться около ста лет.
Химический состав ДНК и её макромолекулярная организация. Типы спиралей ДНК. Молекулярные механизмы рекомбинации, репликации и репарации ДНК. Понятие о нуклеазах и полимеразах. Репликация ДНК как условие передачи генетической информации потомкам. Общая характеристика процесса репликации. Действия, происходящие в вилке репликации. Репликация теломеров, теломераза. Значение недорепликации конечных фрагментов хромосом в механизме старения. Системы исправления ошибок репликации. Корректорские свойства ДНК-полимераз. Механизмы репарации поврежденной ДНК. Понятие о заболеваниях репарации ДНК. Молекулярные механизмы общей генетической рекомбинации. Сайт-специфическая рекомбинация. Генная конверсия.
В 1865г. Грегор Мендель открыл гены, а его современник Фридрих Мишер в 1869г. открыл нуклеиновые кислоты (в ядрах клеток гноя и сперматозоидов лосося). Однако долго еще эти открытия не связывали друг с другом, долго еще структуру и природу вещества наследственности не знали. Генетическая роль НК была установлена после открытия и объяснения явлений трансформации (1928, Ф.Гриффитс; 1944, О. Эвери), трансдукции (1951, Ледерберг, Циндер) и размножения бактериофагов (1951, А. Херши, М. Чейз).
Трансформация, трансдукция и размножение бактериофагов убедительно доказали генетическую роль ДНК. У РНК - содержащих вирусов (СПИДа, гепатита В, гриппа, ВТМ, лейкоза мышей и др.) эту роль выполняет РНК.
Строение нуклеиновых кислот. НК — биополимеры, участвующие в хранении и передаче генетической информации. Мономеры НК — нуклеотиды, состоящие из азотистого основания, моносахарида и одной или нескольких фосфатных групп. В составе НК все нуклеотиды являются монофосфатами. Нуклеотид без фосфатной группы называется нуклеозидом. Сахар, входящий в состав НК, представляет собой D-изомер и β-аномер рибозы или 2-дезоксирибозы. Нуклеотиды, содержащие рибозу, называются рибонуклеотидами и являются мономерами РНК, а нуклеотиды — производные дезоксирибозы, являются дезоксирибонуклеотидами, и из них состоит ДНК. Азотистые основания бывают двух типов: пурины — аденин, гуанин и пиримидины — цитозин, тимин, урацил. В состав РНК и ДНК входят аденин, гуанин, цитозин; урацил встречается только в РНК, а тимин только в ДНК.
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 891; Нарушение авторских прав?; Мы поможем в написании вашей работы!