КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Организация генома эукариот. 1 страница
|
|
|
|
Организменный уровень генетической информации представлен геномом и генотипом. Геном видоспецифичен и представляет тот необходимый набор генов, который обеспечивает формирование видовых характеристик организмов в онтогенезе.
При половом размножении в процессе оплодотворения объединяются геномы двух родительских гамет, образуя генотип нового организма. Все соматические клетки такого организма имеют двойной набор генов, полученных от обеих родителей.
Генотип человека сформировался эволюционно. Общая эволюция генотипа эукариот связана с прогрессивным увеличением количества ДНК. Среди возможных механизмов увеличения генома выделяют полиплоидизацию и амплификацию.
Полиплоидия (увеличение количества ДНК и хромосом, кратное гаплоидному набору) увеличивает дозу всех генов и образует избыток генетического материала, который затем изменяется в результате те мутаций и отбора. В процессе эволюции полиплоидизация сопровождалась переходом в гаплоидное состояние.
Второй механизм – амплификация (образование копий участков ДНК). Определенное значение в увеличении и преобразовании генома имели хромосомные перестройки (делеции, дупликации, транслокации).
Современные представления о геноме человека. Организация генома каждого эукариотического вида представляет собой последовательную иерархию элементов: нуклеотидов, кодонов, доменов, генов с межгенными участками, сложных генов, плеч хромосом, хромосом, гаплоидного набора вместе с внехромосомной и внеядерной ДНК. В эволюционном преобразовании генома каждый из этих иерархических уровней мог вести себя совершенно дискретно (изменяясь, комбинируясь с другими и т.д.).
Наши представления о геноме человека — обширная область генетики человека, включающая, по меньшей мере, понятия «инвентаризации» генов, групп сцепления, картирования генов (локализация), секвенирования всей ДНК (генов, их мутаций и хромосом в целом), мейотических преобразований, функционирования отдельных генов и их взаимодействий, интеграции структуры и функции генома в целом. На решении всех этих вопросов была сосредоточена обширная многолетняя международная программа «Геном человека» (с 1990 по 2003 г.). Главным направлением работ были последовательное секвенирование участков генома и их «состыковка». Успешные разработки в этой области придали программе клинико-генетический аспект (табл.9).
Табл.9. Клинические приложения сведений о геноме человека
| Этапы изучения наследственной лезни | Клинические приложения |
| Регистрация болезни как наследственной формы Локализация гена в хромосоме Выделение гена Определение дефекта гена Обнаружение первичного продукта гена | Медико-генетическое консультирование Дифференциальная диагностика на основе анализа сцепления генов Генотерапия Диагностика(ДНК-специфическая) Диагностика (биохимическая). Улучшение лечения на основе понимания патогенеза |
Систематическое изучение генома человека фактически началось с применения менделевского анализа наследственных признаков человека (начало XX века). Генеалогический метод вошел тогда в широкую практику, и шаг за шагом стал накапливаться материал по «инвентаризации» дискретных наследственных признаков человека, но этот процесс постепенно замедлялся (за 50 лет было открыто не более 400 менделирующих признаков и 4 группы сцепления), возможности клинико-генеалогического метода в чистом виде были исчерпаны.
Бурный прогресс цитогенетики человека, биохимической генетики и особенно генетики соматических клеток в 60-х годах в комплексе с генеалогическим подходом поставил изучение генома человека на новые теоретические основы и высокий методический уровень. Обнаружение новых менделирующих признаков человека стало быстро продвигаться, особенно на биохимическом и иммунологическом уровне, появились возможности изучения сцепления и локализации генов.
Особый импульс изучению генома человека придали молекулярно-генетические методы, или технология генной инженерии (70-е годы). Процесс познания генома углубился до выделения гена в чистом виде и его секвенирования.
В отличие от классической, в новой генетике изменился подход к анализу генов. В классической генетике последовательность была следующей: идентификация менделируюшего признака - локализация гена в хромосоме (или группе сцепления) - первичный продукт гена - ген. В современной генетике стал возможным и обратный подход: выделение гена - секвенирование - первичный продукт, в связи с чем был введён новый термин для определения такого направления исследований: «обратная генетика» или «генетика наоборот».
Продолжаются совершенствование молекулярно-генетических методов и, что не менее важно, их автоматизация. В США и Великобритании были разработаны и внедрены автоматические приборы по секвенированию геномов. Их назвали геномотронами. В них осуществляется до 100 000 полимеразных реакций в час. Это означает, что в течение недели может быть просеквенирован участок (или участки) длиной в несколько миллионов пар нуклеотидов.
Большую роль в расшифровке генома человека играют вычислительная техника и информационные системы. Благодаря им решаются вопросы накопления информации (базы данных) из разных источников, хранения её и оперативного использования исследователями из разных стран.
Характеристика генома человека. Структура генома. Геномом (от слов ген + хромосома) называется совокупность всей наследственной информации организма (всех генов и межгенных последовательностей нуклеотидов). Размер генома человека составляет 3 миллиарда пар оснований. Каждая из 23 пар хромосом содержит отдельную линейную двунитевую молекулу ДНК. Размер ДНК в самой большой хромосоме 1 (хромосомы нумеруют по размеру) - 250 миллионов пар нуклеотидов, а в самой маленькой - 47 миллионов.
В каждой клетке человека около 22-25 тысяч пар генов, но точное их число пока не известно. В генах записана информация о структуре молекул РНК: матричной (кодирующей белки), рибосомной, транспортной и некоторых других видов так называемой некодирующей РНК.
Средний размер гена в хромосоме составляет около 50 тысяч пар нуклеотидов. Самые короткие гены содержат всего два десятка букв-нуклеотидов, например, гены эндорфинов - белков, вызывающих ощущение удовольствия. Гены интерферонов - белков, защищающих человека от вирусных инфекций, имеют размер около 700 нуклеотидов. Самый длинный ген, кодирующий один из белков мышц - миодистрофин, содержит 2,5 миллиона букв.
У примитивных организмов, таких как бактерии, гены занимают около 80-90% всей ДНК. У человека на гены приходится, по-видимому, не более 5% нуклеотидных последовательностей. Остальную ДНК раньше называли избыточной, но со временем стало ясно, что она выполняет важные функции, в том числе содержит информацию о том, как, в каком порядке должны включаться гены. Около трети генома приходится на повторяющиеся последовательности различной длины.
В начале и в конце гена находятся регуляторные последовательности, которые определяют, в каких тканях, на каких этапах развития и при каких внешних или внутренних (например, гормональных) сигналах будет работать данный ген.
Регуляторные последовательности находятся не только рядом с генами, но и в участках, содержащих так называемую реторовирусную ДНК - остатки ретровирусных геномов, которые когда-то встроились в геном человека и переходят в его составе из поколения в поколение. Ретровирусы принадлежат к широкой группе генетических элементов, реплицирующихся с помощью обратной транскрипции. Некоторые ретровирусы не связаны с какой-либо болезнью, тогда как другие очень патогенны, такие как вирус гепатита В и вирус иммунодефицита человека. Ретровирусы инфицируют разнообразные виды позвоночных, от рыб до человека.
Во время репликации ретровирус копирует свой РНК-геном в ДНК, используя кодируемый вирусным геномом фермент - обратную транкриптазу (ревертазу). Вирусная ДНК встраивается в хозяйские хромосомы с помощью другого вирусного фермента - интегразы. Если вирусный геном встраивается в хозяйские гены, это блокирует работу гена. Если участок встраивания находится рядом с геном, то регуляторные элементы вируса могут влиять на работу клеточных генов. Встраивание «чужих» регуляторных последовательностей рядом с генами, отвечающими за чередование фаз деления и роста клетки, приводит к перерождению клетки в раковую.
При встраивании генома вируса в зародышевую линию клеток вирусная ДНК наследуется как менделирующий признак. Большинство вирусных последовательностей встроились в геном предков человека десятки миллионов лет назад. За прошедшее время в них накопилось множество мутаций и они утратили свою патогенность. Часть из них сохранила способность «прыгать» по геному, перенося регуляторные элементы. Эндогенные ретровирусы составляют около 3% ДНК человека.
Гены человека (также как и других эукариотических организмов) имеют сложную структуру. После синтеза РНК некоторые ее участки (их называют вставочными последовательностями или интронами) вырезаются, а оставшиеся (их называют экзонами) сшиваются в единую цепь, содержащую белок-кодирующую последовательность и сигналы регуляции трансляции. Интрон-экзонная структура генов достаточно сложна.
Экзоны могут соединяться в различных сочетаниях, благодаря чему один ген может определять синтез нескольких десятков различающихся своей аминокислотной последовательностью белков. Различия интрон-экзонной структуры зрелой мРНК могут определять интенсивность синтеза одного и того же белка в разных тканях или на разных этапах онтогенеза.
Большинство генов в каждой клетке «молчит». Набор активных генов различается в зависимости от типа ткани, периода развития организма, полученных внешних или внутренних сигналов. Можно сказать, что в каждой клетке «звучит» свой аккорд генов, определяя спектр синтезируемых мРНК, кодируемых ими белков и, соответственно, свойства клетки.
В каждой клетке (кроме эритроцитов, у которых отсутствует ядро) работают гены, кодирующие ферменты репликации и репарации ДНК, транскрипции, компоненты аппарата трансляции (рибосомные белки, рРНК, тРНК, аминоацилсинтетазы и др. ферменты), ферменты синтеза АТФ и другие компоненты, необходимые для ведения «домашнего хозяйства» клетки. Заведуют «домашним хозяйством» около одной пятой всех генов.
ДНК-уровень. Общее количество ДНК в соматической клетке составляет 6,4 х109 пар нуклеотидов, следовательно, гаплоидный набор состоит из 3,2х109 пар нуклеотидов. Основное количество ДНК локализовано в хромосомах (95%). Внехромосомная часть генома человека — ДНК митохондрий (5%). Совсем небольшое количество составляют отдельные кольцевые молекулы ДНК в ядре и цитоплазме.
По способам организации нуклеотидов и функций сегментов ДНК можно выделить следующие фрагменты (рис.29): 1) гены мРНК (структурные гены); 2) гены рРНК; 3) гены тРНК; 4) сателлитная ДНК; 5) спейсерная ДНК.
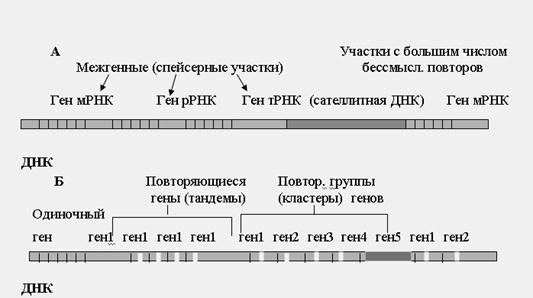
Рис.29. Структурная организация нуклеотидных последовательностей (генов) в ДНК
Структурные гены (тысячи разновидностей) несут информацию о структуре определенных полипептидов. С этих участков ДНК транскрибируется мРНК, которая направляет синтез белков. Гены рРНК (несколько разновидностей) содержат информацию о структуре рибосомальных РНК и обуславливают их синтез. Гены тРНК (более 30 разновидностей) несут информацию о транспортных РНК. Сателлитная ДНК представлена большим числом повторяющихся групп нуклеотидов в разных участках ДНК, которые не имеют смысла и не транскрибируются. Значение сателлитной ДНК полностью пока не известно. Спейсерная ДНК разделяет между собой гены, она не транскрибируется. Роль этих участков до конца не выяснена.
Разные структурные гены имеют особенности организации (рис.29 Б). Например, повторяющиеся гены – один и тот же ген многократно повторяется (много сотен раз), не отделяясь друг от друга, образуя тандемы (например, гены рРНК). Кластеры генов – это группы различных структурных генов на определенном участке хромосомы, объединенных общими функциями.
Например, кластеры пяти разных гистонов повторяются по 10-20 раз. Одиночные гены среди сателлитной ДНК обычно имеют регуляторное или усиливающее действие на структурные гены, например, энхансеры.
Внехромосомные и кольцевые молекулы ДНК обнаруживаются в цитоплазме и ядре. У человека они изучены ещё недостаточно. В строгом смысле они являются не составными элементами генома, а его продуктом. Их размер колеблется от 150 до 20 000 пар нуклеотидов. Являются эти молекулы продуктом фрагментации хромосомной ДНК в клетке или образуются за счёт других генетических процессов (гомологичная рекомбинация, обратная транскрипция), пока неясно. Исследованные к настоящему времени у млекопитающих большие кольцевые молекулы ДНК размером от 150 до 900 000 пар нуклеотидов, локализованные только в ядрах, представляют собой амплифицированные участки онкогенов или генов устойчивости к ядам и антиметаболитам. С этими молекулами предположительно связывают устойчивость клеток к лекарствам и способность клеток к неограниченному росту. Их происхождение объясняют делециями соответствующих областей хромосом.
Хромосомная ДНК подразделяется на две группы участков: с уникальной последовательностью пар нуклеотидов и с повторяющимися последовательностями. Из общей массы ДНК в клетке примерно 50% ДНК с уникальными последовательностями и 50% — с повторяющимися.
Часть ДНК, кодирующая белки составляет всего 3—5%. Что делает «покоящаяся» часть генома, неизвестно. Однако трудно предположить, что она не имеет функций.
Полиморфизм. Любые изменения в структуре ДНК (в хромосомах или митохондриях) ведут к генетическому полиморфизму. Эти изменения могут быть качественными, если они обусловлены заменой или потерей нуклеотидов, либо количественными, если в определённом локусе варьирует число нуклеотидных повторов различной протяжённости. И те и другие варианты генетического полиморфизма встречаются как в смысловых (внутриэкзонных), так и в несмысловых (внегенных или интронных) последовательностях молекулы ДНК.
Главной формой генетического полиморфизма является однонуклеотидный полиморфизм (ОНП). Под этим термином понимают варианты последовательностей ДНК у разных людей с вовлечением одной пары нуклеотидов (рис.30).
| Индивид | Последовательность |
| AG-A-GTT-C-TGC-T-CG AG-G-GTT-A-TGC-G-G | |
| CGTT-C-GG-G-ATC-С CGTT-A-GG-A-ATC-T | |
| TCTT-T-GA-C-ACTC TCTT-A-GA-G-ACTC |
Рис.30. Примеры нуклеотидного полиморфизма у двух индивидов
На данном рисунке представлены три фрагмента последовательностей от двух индивидов. В прямоугольниках выделены однонуклеотидные различия в геномных последовательностях. ОНП — наиболее общий источник вариаций между людьми. Эти вариации встречаются на протяжении всей ДНК (в экзонах, интронах, межгенных промежутках, повторах) и отражают прошлые мутации.
Секвенированием геномов или их частей разных людей установлено, что однонуклеотидные различия обнаруживаются на протяжении 1000—2000 нуклеотидной длины. Это означает, что на всю длину генома (3,2 млрд пар нуклеотидов) должно быть 1,6—3,2 млн ОНП. К 2001 г. идентифицировано и картировано 1,42 млн ОНП. Расчёты показывают, что два человека на 99,9% идентичны по нуклеотидным последовательностям, т.е. только 0,1% различий по одному нуклеотиду создаёт такие огромные индивидуальные фенотипические вариации, которые легко видеть в любой группе индивидов.
Предполагают, что различия по одному основанию между определёнными отрезками геномов лежат не только в основе генных болезней (миссенс-мутации), но и в основе чувствительности к возбудителям или защиты от них, в основе приспособительных реакций и наследственного предрасположения к мультифакториальным болезням. К началу 2001 г. идентифицировано 60 000 ОНП в генах (их называют кодирующими ОНП). Это означает, что в генных последовательностях один кодирующий ОНП встречается в пределах 1080 пар нуклеотидов. Хотя информация об ОНП ещё не полная (основные сведения получены в последние 2 года), уже известно, что 93% генов содержат ОНП. Главное использование карты ОНП — выяснение вклада индивидуальных генов в болезни комплексной (многофакторной) и полигенной природы. Сравнение частот определенных типов ОНП у пациентов и в контрольных группах позволяет идентифицировать ОНП, с которыми ассоциируется заболевание. Несмотря на большие перспективы, которые открываются для объяснения заболеваний человека с пониманием природы и размаха ОНП, необходимо помнить об опасности геномомании. Гены и геномы действуют не в вакууме. Среда не менее важна для биологии человека, чем гены. Карты ОНП при правильном использовании позволяют лучше понять роль природы (генотипа) и среды в широком понимании в развитии человека в целом и патологии в частности.
Генетические карты. Составной частью сведений о геноме человека наряду с нуклеотидной последовательностью являются генетические карты хромосом, т.е. схемы, описывающие порядок расположения генов и других генетических элементов на хромосоме с указанием расстояния между ними. Генетическое расстояние измеряется по частоте рекомбинации между гомологичными хромосомами и выражается в сантиморганидах (сМ). Одна сМ соответствует частоте рекомбинации, равной 1%. Длина всего генома человека равна примерно 3000-3500 сМ.
 Изучение групп сцепления и составление карт хромосом первоначально основывались на анализе «расщепления» фенотипов в потомстве формально-генетическими методами. Применение молекулярно-генетических методов значительно ускорило картирование генов, а секвенирование генома позволяет составить полные генетические карты для всех хромосом. На рис.31 представлена в качестве примера карта хромосомы 3 по генам, патологические мутации в которых ведут к наследственным болезням. Такие карты называют патологической анатомией генома человека. Следует отметить, что это далеко не полная карта, она постоянно уточняется.
Изучение групп сцепления и составление карт хромосом первоначально основывались на анализе «расщепления» фенотипов в потомстве формально-генетическими методами. Применение молекулярно-генетических методов значительно ускорило картирование генов, а секвенирование генома позволяет составить полные генетические карты для всех хромосом. На рис.31 представлена в качестве примера карта хромосомы 3 по генам, патологические мутации в которых ведут к наследственным болезням. Такие карты называют патологической анатомией генома человека. Следует отметить, что это далеко не полная карта, она постоянно уточняется.
Рис.31. Патологическая анатомия хромосомы 3
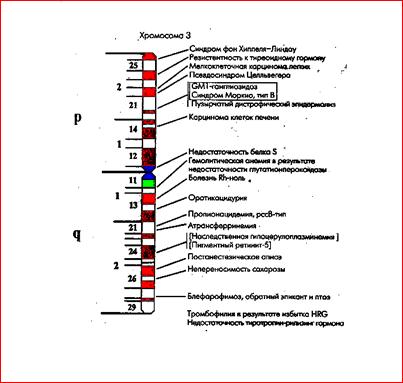 |
Знание генетических карт необходимо в разных разделах медицинской генетики: для диагностики болезней методом сцепления; оценки патологических эффектов хромосомных транслокаций; решения вопросов эволюционной и популяционной генетики.
Гены, кодирующие полипептиды, РНК. Гены, кодирующие белки, как правило, содержат кодирующие области (экзоны), прерываемые одним или более нитронами. Однако некоторые гены (например, гистоновые или кодирующие интерфероны) не имеют вставочных последовательностей. Число таких последовательностей сильно варьирует от гена к гену, а количество ДНК, приходящееся на долю интронов, во много раз превышает количество ДНК кодирующих областей — у некоторых организмов в десятки раз. Как правило, нуклеотидные последовательности аналогичных экзонов, относящихся к паралогичным генам в данном геноме или к ортологичным генам в геноме разных видов, более консервативны, чем нуклеотидные последовательности соответствующих интронов. Положение интронов, как правило, фиксировано, а по длине и составу они варьируют.
Все известные гены, кодирующие белки, транскрибируются РНК-полимеразой II и поэтому часто имеют сходные промоторы и сигналы полиаденилирования. Но многие такие гены связаны с более специфичными регуляторными последовательностями, которые опосредуют действие гормональных, средовых или онтогенетических факторов.
В отличие от генов, кодирующих РНК, полипептидные гены представлены в геноме в единственном числе, однако при этом геном часто содержит сегменты, гомологичные данному специфическому гену. Таким образом, однокопийный ген может входить в состав семейства близкородственных последовательностей (например, в семейство генов гормона роста). Члены такого семейства могут кодировать незначительно различающиеся белки (например, изозимы). Однако они могут иметь разные регуляторные сигналы, ответственные за экспрессию генов в разных тканях или на разных стадиях развития (как в случае генов гормона роста или плацентарного лактогена). Равным образом членами семейства могут быть и псевдогены. Условились, что два неаллельных гена считаются идентичными, если они кодируют фактически одинаковые белки и находятся под общим контролем.
Мультигенные семейства. Семейства генов обычно кодируют белки, богато представленные в клетке. В своем большинстве гены активны. По критерию соответствия последовательностей специфичному зонду обнаруживаются много генов кодирующих синтез интерферона, актина, тубулина... Внутри семейства генов некоторые его члены могут повторяться чаще, чем другие. Например: существует 8 генов для а-типа интерферона человека, и только один β-типа.
Многие гены могут быть объединены в кластеры или рассеяны по геному. Общий план строения глобиновых генов консервативен. Гены-интерфероны имеют сходную структуру, для которой характерно отсутствие интронов. Гены актина имеют прерывистую структуру. У этих генов участки, кодирующие белок, обладают высокой степенью гомологии. Таким образом, если активные гены произошли от общего гена-предка, расположение экзонов и интронов претерпело существенные изменения. Функционирующие гены могут иметь прерывистое и непрерывное строение и изменения в расположении интронов не обязательно влияют на активность генов. Гены, кодирующие одинаковые или близкие белки, необязательно должны быть организованы в виде тандема, но могут быть рассеяны по геному в виде отдельных индивидуальных генов или малых кластеров. Повторение последовательностей приводит к увеличению количества ДНК, кодирующей определенную функцию. Возможно существование нескольких активных генов, кодирующих либо один и тот же белок, либо неожиданные его варианты.
Когда копии генов объединены в кластеры, расстояние между ними может быть значительным, что может увеличивать общее количество ДНК, ответственное за осуществление данной функции: основная часть функции кодируется несколькими генами, а не уникальным геном; значительное количество ДНК не несет кодирующей функции.
Однокопийные гены могут также принадлежать большому семейству отдаленно родственных последовательностей. Несмотря на структурное сходство, гены такого надсемейства кодируют совершенно разные белки (как, например, уже упомянутые гены пролактина и гормона роста). Распределение отдельных генов по надсемействам не всегда удается провести вполне четко. Часто сходство нуклеотидных последовательностей членов варьирует от 95% и более до 50% и менее. В жестких условиях отжига дуплексы образуют только сегменты ДНК с гомологией не менее 80%, поэтому неблизкородственные семейства можно сравнивать, лишь зная их нуклеотидную последовательность. Более того, сходство между двумя белками не находится в простой зависимости от процентного сходства между соответствующими генами. Вследствие вырожденности генетического кода значительные изменения в третьей позиции кодонов мало влияют на тип кодируемой аминокислоты, а такие же изменения в первой или во второй позициях приводят к существенным изменениям в структуре белка.
Помимо генов, принадлежащих мультигенным семействам или надсемействам, имеются и уникальные гены (например, ген тиреоглобулина человека и ген актина дрожжей). Каждому мультигенному семейству генов, кодирующих белки, присущи особые свойства, характерные для данного вида.
Актиновые гены: мулътигенное консервативное семейство. Актин участвует в самых разных типах клеточного движения, в частности он обеспечивает клеточную подвижность и мышечные сокращения. Соответственно эукариоты имеют несколько разных типов актина и кодирующих их генов. Одни из этих генов организованы в кластеры, другие диспергированы. В ходе эволюции актиновых генов в них встраивались новые интроны. Каждое такое событие приводило бы к смещению соседних последовательностей и к сдвигу рамки считывания. Можно предположить обратное: большинство актиновых генов содержали интроны во всех тех положениях, где они находятся и в современных генах, но при эволюции разных таксонов утрачивались различные интроны.
Актиновые гены в основном рассеяны по хромосомам. У мыши, например, гены а-актина скелетных мышц и сердечной мышцы локализованы в хромосомах 3 и 17 соответственно, а гены цитоплазматического β-актина ~ в хромосоме 5.
У большинства изученных организмов число актиновых генов, по-видимому, превышает число известных актиновых белков. Хотя не исключено, что какие-то из этих «экстра»- последовательностей кодируют не обнаруженные пока формы актина, большинство из них, вероятно, являются псевдогенами, в том числе процессированными. У человека по меньшей мере две из двадцати актиноподобных последовательностей представляют собой процессированные псевдогены β-актина; они не содержат интронов и несут в кодирующих областях различные мутации.
Тубулиновые гены: мультигенное семейство, которое включает гены двух разных субъединиц гетеродимерного белка. Микротрубочки участвуют во многих процессах, протекающих во всех эукариотических клетках: мейозе, митозе, клеточном движении и секреции. Поэтому не удивительно, что структура тубулина-белка, из которого состоят микротрубочки, равно как и нуклеотидная последовательность гена, кодирующего тубулин, одинаковы у всех эукариот. кДНК, синтезированные на тубулиновой мРНК курицы, гибридизуются с тубулиновыми генами таких отдаленных организмов, как дрожжи и млекопитающие.
Тубулин - это гетеродимер, состоящий из двух полипептидов: а и β. а- и β-субъединицы содержат 450-451 и 445 аминокислотных остатка соответственно и гомологичны примерно на 40%. Гены обеих субъединиц относятся к одному мультигенному суперсемейству, хотя довольно сильно различаются и не гибридизуются. Как правило, семейства тубулиновых генов у каждого вида эукариот кодируют разные изотипы а- и β-субъединиц. Аминокислотные последовательности различных а- или β-субъединиц, кодируемых этими генами, обычно различаются лишь незначительно (менее чем на 10%), и различия касаются в основном карбоксильных концов молекул. По-видимому, вариации в структуре тубулинов связаны с тем, что немного различаются те микротрубочки, которые они образуют. Последние в свою очередь специфичны в отношении клеточных процессов, типов клеток и стадий развития. Таким образом, разные а- и β-изотипы могут соответствовать разным типам микротрубочек, выполняющих определенные функции, хотя такая специализация и не абсолютна. Дальнейшие изменения в структуре тубулина происходят уже после транскрипции и тоже способствуют функциональной специализации микротрубочек. О такой специализации свидетельствует высокая консервативность специфических изотипов у позвоночных. Например, у различных позвоночных преобладающий в нервной ткани β-тубулин имеет на карбоксильном конце совершенно одинаковые последовательности.
Суперсемейства генов и их продукты. Суперсемейство глобиновых генов. Гемоглобины позвоночных — это гетеротетрамеры, содержащие по два а- и β-полипептида. В геноме всех позвоночных содержится множество генов и псевдогенов а- и β-цепей- членов суперсемейства, которое включает кодирующие последовательности генов глобинов беспозвоночных, миоглобина позвоночных и леггемоглобина растений. Все эти белки содержат гем и обратимо связываются с кислородом. Глобиновые мРНК и соответствующие гены были среди первых объектов, которые исследовались методами рекомбинантных ДНК. На долю гемоглобина приходится более 90% всех растворимых белков эритроцитов, а на долю глобиновой мРНК — большая часть мРНК ретикулоцитов и ядерных еритроцитов. Ни в каких других клетках глобиновые гены в заметном количестве не транскрибируются. Возможность получения относительно чистой глобиновой мРНК, наличие огромного числа уже известных мутаций в генах глобина человека, а также обширные данные о свойствах глобиновых белков послужили стимулом к молекулярным исследованиям этих генов. Удалось получить первые данные об интронах в клеточных генах и о свойствах промоторов для РНК-полимеразы II. Была установлена нуклеотидная последовательность различных аллелей глобиновых генов у человека и других видов, что послужило основой для создания многочисленных теорий эволюции глобинов.
Как правило, у млекопитающих имеется множество глобиновых генов и псевдогенов. У человека β-глобиновые гены объединены в кластер длиной 65 т. п. н., расположенный в хромосоме II, а а-глобиновые гены — в кластер длиной 25 т. п.н. в хромосоме 16. Кластер β-глобиновых генов содержит пять генов, а ặ-глобиновый кластер — три. Оба кластера включают также по нескольку псевдогенов.
У человека и других млекопитающих различные глобиновые гены экспрессируются на разных стадиях развития организма. У многих видов, в том числе и у человека, расположение генов в а- и β-кластерах соответствует тому порядку, в котором они экспрессируются во время развития организма.
Интерфероновые гены. В ответ на разнообразные внешние воздействия клетки многих позвоночных секретируют полипептиды, называемые интерферонами. Например, в результате вирусной инфекции и попадания в клетку двухцепочечной РНК в лейкоцитах индуцируется синтез интерферонов группы ặ, а в фибробластах-интерферонов группы β (IFN-а, I IFN-β, 1 IFN-γ).
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 1982; Нарушение авторских прав?; Мы поможем в написании вашей работы!