КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Материнская часть плаценты образуется за счет основной отпадающей оболочки (части) миометрия 1 страница
|
|
|
|
(слизистой оболочки матки). Па раллельно происходят изменения в хорионе. Эти изменения обеспечивают выполнение 2-х задач: снабжение растущего зародыша веществами, необходимыми для его развития, предохранение матки от чрезмерного проникновения хориона.
Комплекс этих изменений приводит к превращению слизистой оболочки матки в децидуальную ткань, наличие которой является главным критерием для деления плацентарных животных на группу адецидуатов, слизистая матки которых не изменяется, и децидуатов, эндометрий которых подвержен децидуальным изменениям.
В децидуальную ткань превращается соединительная ткань стромы слизистой оболочки матки. В соединительной ткани в большом количестве образуются децидуальные клетки. В их цитоплазме появляются многочисленные зерна гликогена и липиды. Значительно увеличивается гидратация слизистой оболочки, становятся менее различимы ретикулярные и коллагеновые волокна, значительно увеличивается просвет сосудов. Эти изменения приводят к тому, что слизистая оболочка матки становится толстой, сочной и рыхлой.
Физиологическое значение децидуальной реакции заключается в том, что для плода она обеспечивает питание и защиту. Трофическое значение ее проявляется, главным образом, в ранней гистотрофной фазе питания плодного яйца. Зигота перед имплантацией питается, в основном, продуктами слизистой и, прежде всего, секретом децидуальных желез. Далее, главным источником трофного питания являются продукты цитолиза децидуальных клеток.
Защитная функция децидуальной ткани проявляется в ее исключительной фагоцитарной активности. Благодаря этой способности она связывает и разрушает токсины, бактерии, проникающие из организма матери к плоду.
Для матери защитная роль децидуальной системы состоит в торможении ею внедрения трофобласта. Это осуществляется благодаря наличию в Децидуальной ткани протеолитических ферментов, которые во второй половине беременности сохраняются только в более глубоких слоях слизистой.
К защитным факторам децидуальной ткани относится наличие в ней в значительных количествах фибринобластических (способствующих свертыванию крови) п. фибринолитических (усиливающих процесс расплавления тромбов) факторов. На более^оздних этапах беременности эта ткань замещается фибрингиалиноподоб- ной массой.
Фибриноид образуется в месте непосредственного контакта синцитотрофобласта и децидуальных клеток.
Плацента выполняет ряд функций: трофическую, дыхательную, выделительную, барьерную, эндокринную.
1. Трофическая: из крови матери в кровь плода поступают различные питательные вещества (аминокислоты, глюкоза, нуклеотиды, витамины), необходимые для.нормального развития зародыша.
2. Дыхательная: кровь плода богата углекислотой, которая легко и быстро диффундирует через плаценту. Кислород диффундирует не так легко и для обеспечения плода достаточным количеством кислорода напряжение его в тканях матери должно быть значительно выше, чем по другую сторону плацентарного барьера. Установлению этого градиента способствует также отличие гемоглобина плода от гемоглобина взрослого организма по сродству к кислороду. При одном и том же напряжении кислорода гемоглобин плода способен связывать больше кислорода, чем гемоглобин матери.
3. Выделительная функция плаценты заключается в выведении продуктов распада (мочевина, аммиак и др.) из крови плода в кровь матери.
Трофическая, дыхательная и выделительная функции плаценты обеспечиваются с помощью организма матери. Поэтому беременная женщина должна соблюдать нормальный режим, не курить, не принимать алкоголь, который беспрепятственно проходит через плаценту и разрушает лервную,- систему плода. Будущая мать должна быть на воздухе, следить за работой своего выделительного аппарата.
4. Барьерная функция. Плацента является барьером, через который не проходят из крови матери в кровь плода многие вещества, которые могут нарушить нормальное течение эмбриогенеза. Ни один микроб, кроме бледной спирохеты, вызывающей сифилис, не проникает через плаценту. Плацента защшвдет организм матери от антигенов плода, препятствует развитию иммунобиологического конфликта.
5. Эндокринная: плацента является временной эндокринной железой, которая обеспечивает нормальное течение беременности. Один из первых гормонов, который был выделен из плаценты — это гонадотропный гормон. Максимум его секреции достигается к 8—9 неделе, а затем содержание его снижается. Вырабатывают его клетки Лангганса. Планцета вырабатывает АКТГ (при беременности улучшается состояние больных Адисоновой болезнью, заболеваниями соединительной ткани — коллагенозами), тиреот- ропный гормон, меланОфорный, лютеотропный, эстрогены, прогестерон, андрогены, кортикоиды. Вьфабатываемый плацентой релаксан вызывает расслабление лонного сочленения при беременности. В плаценте вырабатывается гистамин, под его влиянием расширяются капилляры, а гладкая мускулатура — сокращается. Гистамины разрушаются гистаминазами, которые вырабатываются плацентой в большом количестве. Когда плацента стареет, количество гистаминазы уменьшается и гистамины приводят мышцы матки к родовой деятельности. Образующийся в плаценте ацетилхолин оказывает то же действие, что и гистамин.
Таким образом, плацента обеспечивает нормальное развитие
зародыша. После рождения плода материнская и дочерняя части плаценты отторгаются из матки. Это получило название рождение последа.
ТЕОРЕТИЧЕСКИЕ ОСНОВЫ ПОНИМАНИЯ ЭМБРИОГЕНЕЗА
В предыдущих разделах. были описаны основные этапы эмбриогенеза. Как известно, филогенез дает историю вида, онтогенез 45- историю индивида. В свою очередь онтогенез, описывающий историю индивида от момента зарождения до наступления физиологической смерти, может быть подразделен на эмбриональный и постэмбриональный период. Эмбриология как наука изучает эмбриональный период онтогенеза. У плацентарных, к которым относится и человек, эмбриональный период захватывает время от момента оплодотворения до рождения, т. е. до момента изгнания плода из матки.
Рассматривая многочисленные работы, выполненные по эмбриологии в течение нескольких столетий, можно юс подразделить на описательные или феноменологические и на экспериментальные, стремящиеся вскрыть причины и механизмы, обусловливающие сложные и не до донца выясненные процессы эмбриогенеза.
Описание эмбриогенеза позволяет выделить в нем стадии, процессы, элементы и элементарные акты. Под стадией следует понимать этап развития зародыша, коренным образом отличающийся от предыдущего в силу появления новых свойств и структур. Исходя из этого определения можно выделить следующие стадии эмбриогенеза: зигота, бластула, гаструла, нейрула, тканевое и органное строение.
.Переход от одной стадии к следующей обеспечивается с помощью определенных эмбриональных процессов.
Так, появлению стадии зиготы, предшествует процесс прогенеза, сущностью которого является образование и созревание половых клеток, их перенос навстречу друг другу с последующим слиянием. Оплодотворение является процессом, приводящим к появлению зиготы. Биологическая сущность оплодотворения состоит в том, что две половые клетки с гаплоидным набором хромосом, не способные раздельно к прогрессивному развитию и обреченные на быструю гибель, при встрече сливаются и образуют диплоидную одноклеточную стадию организма — стадию зиготы с обширной и реализуемой программой прогрессивного развития.
Процесс, с помощью которого зигота переходит в следующую стадию развития — бластулу, получил название дробления. Дробление совершается, используя известный механизм митотическо- го деления, но с рядом важных особенностей. Во-первых, при дроблении интерфаза резко укорочена. Она представлена, по сути, периодом, в котором протекает репликация. Во-вторых, при дроб-
момента оплодотворения до рождения на свет (у живородящих организмов) или до вылупливания из яйцевых оболочек (у яйцекладущих животных)». (А. Г. Кнорре, 1914—1982).
Предметом изучения эмбриологии является также прогенез — предзародышевый период (формирование половых клеток) и по- слезародышевый- (постэмбриональный) или послеродовой (постна- тальный) периоды индивидуального развития. У млекопитающих: некоторые системы органов (например, половая, эндокринная) приобретают дефинитивные, т. е. окончательные, свойственные взрослому состоянию, структуры и функции спустя тот или иной промежуток времени после рождения.
Эмбриология исследует как общие закономерности индивидуального развития, проявляющиеся в развитии всех'многоклеточных животных организмов, так и частные особенности процессов индивидуального развития, характерные для представителей отдельных типов, классов или видов животных.
В связи с этим различают общую эмбриологию, рассматривающую наиболее общие вопросы и наиболее широкие закономерности индивидуального развития, и частную эмбриологию отдельных групп животных.
Значение эмбриологии заключается прежде всего в формировании общих биологических представлений на- материалистической основе. Велико значение эмбриологии как теоретической основы педиатрии, акушерства и гинекологии, тератологии.
МЕТОДЫ ИССЛЕДОВАНИЯ
Для изучения эмбрионального развития животных пользуются различными методами.
Простейший и наиболее древний из них — наблюдение и описание развития зародыша у тех или иных животных или человека. На основе этого метода возникла описательная эмбриология.
Значительно позже в эмбриологических исследованиях стал применяться сравнительный метод, создавший сравнительную эмбриологию. Ее возникновение было обусловлено накоплением фактов о развитии различных видов животных. Применение сравнительного метода открыло возможности для понимания общих закономерностей развития. С помощью сравнительного метода был открыт закон зародышевого сходства К. М. Бэром (1792—1876).
Описательная и сравнительная эмбриология стали фундаментом для развития эволюционной эмбриологии, создателями которой явились А. О. Ковалевский (1840—1901), И. И. Мечников (1845— 1916). Развитие эволюционной эмбриологии привело к открытию биогенетического закона Э. Геккелем (1834—1919), Ф. Мюллером (1821—1897), А. Н, Северцевым (1861—1936).
С развитием и усложнением экспериментальной техники воз
никло новое направление исследований — экспериментальная эмбриология. Основателями ее явились В. Ру (1850—1924), Г. Дриш (1867—1941) и Г. Шпеман (1869—1941). В основе экспериментальной эмбриологии лежит изучение последствий удаления или трансплантация различных частей зародыша, а также воздействия на зародыш или его части разных экзогенных факторов.
Современная эмбриология пользуется всеми видами световой, электронной и сканирующей микроскопии, методами прижизненного наблюдения эмбриогенеза при помощи меток, наносимых на зародыш с помощью изотопов или красителей, методами гистохимии, биохимии, радиологии, иммунологии, а также статистики.
ТЕОРИИ ЭМБРИОЛОГИИ
. Как и любая другая наука, эмбриология на каждом этапе развития была вооружена собственной теорией.
С именем Гиппократа (460—377 год до н. э.) и Аристотеля (384—322 год до н. э.) связывают не только определенные знания в области развития организмов, но и создание первых научных эмбриологических представлений.
Согласно теории Гиппократа, плод образуется через смешение мужского и женского семени. Он считал, что все члены организма 1возникают независимо друг от друга. Ни один из них не возникает раньше или позже другого.
Эта идея вновь возродилась и стала господствовать в течение XVII—XVIII вв. Она получила название теории преформизма. Согласно этой теории каждый зародыш с самого начала является уже вполне сформированным, имеющим все части организма, которому остается только расти. Наиболее крайние и наивные преформисты представляли себе, что каждый организм, в том числе и человеческий, заключает в себе громадное количество поколений зародышей, вложенных друг в друга в теле прародителей со времени сотворения мира.
Между представителями двух направлений преформистов велись горячие споры. Анималькулисты или сперматики считали, что зародыш преформирован в сперматозоиде. Овисты думали, что зародыши в миниатюрном виде находятся не в мужских половых клетках, а в яйце.
Сторонниками префоризма были многие выдающиеся ученые XVII и XVIII века: А. Левенгук (1632—1723), Я. Сваммердам (1637—1680), М. Мальпиги (1628—1694), А. Галлер (1708—1777), Ш. Бонне (1720—1793).
Аристотель впервые сформировал теорию, эпигенеза. Согласно Аристотелю, части зародыша не заложены в невидимо малых размерах в «семенах вещей» и не возникают все одновременно, а растут и развиваются в определенной последовательности. Это
лении имеет место в интерфазе только синтез и-РНК, остальные виды РНК и синтез белка еще не происходят. Отсюда, клетка не способна к росту и поэтому при каждом новом этапе дробления дочерние клетки-бластомеры получают размеры вдвое меньше,, чем материнские.
Если вспомнить положение биогенетического закона и приложить его к дроблению, то этот процесс онтогенеза как бы повторяет в ускоренном виде переход от одноклеточных к многоклеточным формам-, который имел место в филогенезе. '
Следующий процесс, эмбриогенеза — это гаструляция, с помощью которой зародыш из стадии бластулы переходит в стадию гаструлы. В основе этого процесса лежит перемещение клеточных масс, приводящее вначале к появлению двух, а затем и трех зародышевых листков: эктодермы, энтодермы и мезодермы: Причины и механизмы этого перемещения клеток выяснены недостаточно.
. Продолжающееся перемещение клеток тела зародыша — процесс нейруляции, приводит к образованию органов спинного комплекса: нервной трубки, хорды, сомитов, тем самым приводит зародыш на стадию нейрулы. На этой стадий развития зародышевые зачатки заняли «исходные рубежи» и готовы к следующему: процессу — гистогенезу, т. е. к образованию тканей. Параллельно- с^образованием тканей их взаимодействие приводит к появлению новых структур — возникают органы. Процесс образования органов, получил название - органогенеза. Очень часто оба эти процесса объединяют под общим термином морфогенеза.
Процессы морфогенеза продолжаются на протяжении всей жизни человека или животного. В настоящем разделе мы рассмотрим эти процессы в эмбриональном периоде..
Все процессы эмбриогенеза можно условно разделить на элементы. К ним относятся: размножение клеток, рост клеток, образование некоторыми из, них межклеточного промежуточного вещества, перемещение клеток, детерминация и дифференцировка клеток, взаимодействие и гибель клеток.
Размножение клеток как элемент эмбриогенеза представлен дроблением, митозом и мейозом. Основная масса клеток возникает в результате митотического деления. Роль амитоза, как самостоятельного вида деления клеток, в. условиях нормального развития организма в настоящее время пересматривается. Мейоз свойственен только половым клеткам в период их созревания. У большинства видов животных, в том числе и у человека; он протекает в постэмбрйональном периоде.
Размножение клеток и последующий их рост, начиная со стадии гаструлы, генетически запрограммированные процессы, регулируемые действием биологически активных веществ. Рост клеток объясняют преобладанием процессов ассимиляции над
процессами диссимиляции. Уравнение интенсивности этйХ^процес- «сов приводит к остановке роста клеток.
Одну из загадок эмбриогенеза представляют способы и механизмы перемещения клеток. Можно выделить перемещение клеток пластом, как это имеет место при гаструляции и нейруляции; перемещение группами или отдельными клетками, как это имеет место при миграции первичных половых клеток, нейробластов в -состав ганглиев, миобластов из миотомов в места локализации -будущих скелетных мышц. Механизм перемещения отдельных клеток объясняют * с позиций амебоидного и ундулирующего движения. Направление движения определяется хемотаксисом, а •остановка'—механизмом контактного торможения,•• в основе которого лежит прекращение движения при встрече однотипных клеток.,
Прежде чем перейти к рассмотрению механизмов детерминации:и дифференцировки, дадим определение понятию «элементарный акт». В основе всех явлений жизни, как особой формы движения матери, а, Следовательно, процессов, протекающих в клетках, тканях и органах на этапах эмбриогенеза, лежат элементарные -акты морфогенеза.
Под элементарным актом мы понимаем цикл молекулярно-фер- ментативных. процессов, приводящих к образованию макромолекул или сборке надмолекулярных субклеточных структур.
К элементарным актам морфогенеза следует отнести процессы репликации, приводящие к., синтезу молекул ДНК, процессы транскрипции—ксинтезу всех.видов молекул РНК (и-РНК, р-РНК, т-РНК, рег.-РНК), процессы трансляции, приводящие к синтезу молекул белка, а также' процессы самосборки, приводящие к образованию различных мембран и органелл.
При эмбриогенезе появление и становление элементарных актов морфогенеза происходит в определенной последовательности. В зиготе возникшей из изолецитальной яйцеклетки, еще не представлены макромолекулярные синтезы. В период дробления у зародышей этого типа появляются процессы репликации и начинается синтез и-РНК. На стадии бластулы к этим элементарным актам присоединяется синтез т-РНК, на стадии гаструлы — р-РНК наконец, на стадии нейрулы в клетках зародыша представлены все виды макромолекулярных'синтезов и сборки надмолекулярных структур. Тем самым, на стадии нейрулы получили развитие все элементарные акты, обеспечивающие дальнейшее эмбриональное развитие.
Стадия нейрулы является стартовой площадкой для; дальнейших процессов гистогенеза, объединяемых общим термином «морфогенез». Под морфогенезом следует понимать цепь последовательно развивающихся процессов, приводящих к становлению определенных структур и функций.
Исследуя начальные, промежуточные и конечные этапы морфогенеза, нетрудно заметить, что различные по структуре и функциям структуры возникают часто из одинаковых зачатков и отличия между ними нарастают постепенно. Так, нервная ткань и развивающийся из нее спинной и головной мозг возникают из общего с кожей зачатка — эктодермы. Жидкая кровь, костная ткань и жировая ткань у зародыша имеют общего родоначальника в виде мезенхимы. Вместе с тем, клетки крови резко отличаются как от клеток костной, так и жировой ткани по своим размерам, форме, функции, биохимизму, что указывает, что активность генов, определяющих все эти отличия, различна у клеток этих тканей.
Процесс, который при онтогенезе приводит к развитию специализированных по структуре и функции клеток, получил название дифференцировки. Как объяснить, что одинаковые по своему происхождению, структуре и функции клетки мезенхимы выбирают различные направления развития, т. е. дифференцируются по-разному? Для объяснения этого очень важного положения введено понятие о детерминации.
Под детерминацией следует понимать стойкое и передающееся по наследству изменение состояния регуляторных структур клетки, определяющее направление дифференцировки.
Итак, под действием определенных факторов в клетке происходит детерминация, т. е. клетки получают программу развития. Затем наступает дйфференцировка, с помощью которой реализуется программа детерминации.
Понимание механизмов развития, что получило название «механики развития», создавалось работниками экспериментаторов-эмбриологов на протяжении более 100 лет и существенно продвинулось благодаря успехам молекулярной биологии, молекулярной генетики и электронной микроскопии.
Как известно, из зиготы путем дробления возникают бластомеры, из которых в свою очередь развиваются различные клетки зародыша. Решим, во-первых, вопрос, какова роль ядра клеток в процессе дифференцировки клеток. С этой целью рассмотрим результаты опытов Бриггса и Кинга.
Излюбленным объектом для решения вопросов механики развития являются зародыши амфибий, в частности, головастики. Это объясняется доступностью объекта, неприхотливостью, высокой скоростью эмбрионального развития.
Если удалить ядро из зиготы головастика с помощью специальной иглы, т. е. энуклеировать зиготу, то последняя совершит несколько дроблений, но возникшие энуклеированные бластомеры вскоре погибнут и дальнейшее развитие прекратится. Вывод ясен — ядро необходимо для процессов эмбриогенеза.
Эксперимент несколько изменяется: в энуклеированные зигбты будущих головастиков трансплантируются ядра, выделенные из
бластомеров или из клеток зародыша, находящихся на стадии бластулы, ранней гаструлы, поздней гаструлы и нейрулы. Затем ведется наблюдение за судьбой зародыша. В подавляющем проценте случаев энуклеированные зиготы, которым были трансплантированы ядра из клеток бластулы и ранней гаструлы проходят все этапы развития и превращаются в нормальных головастиков. Эти результаты позволяют сделать выводы, что ядра клеток зародыша, находящегося на этих стадиях, по своим свойствам не отличаются от ядер зиготы, т. е. они эквипотентны, а поскольку они обеспечивают различные дифференцировки в процессе эмбриогенеза, то ядра клеток бластулы и ранней гаструлы подобно ядрам зиготы полипотентны.
При трансплантации ядер клеток зародышей, находившихся на стадиях поздней гаструлы и нейрулы, развитие головастиков прекращается из-за гибели на стадии ранней гаструлы. Следовательно, при эмбриогенезе/ ядра клеток зародышей уже на стадии поздней гаструлы претерпевают изменения и теряют способность обеспечивать развитие зиготы до взрослой формы. На стадии поздней гаструлы ядра клеток головастиков уже не экви- и не полипотентны. На основании этих опытов можно сделать вывод, что в процессе эмбриогенеза изменяются свойства ядер клегок: уменьшается их способность обеспечивать развитие клеток в различных направлениях, т. е. они специализируются или иначе дифференцируются.
В отличие от Бриггса и Кинга, Гердон в: более поздних работах обнаружил, что в 1% случаев, т. е. очень, редко, ядра клеток эпителия кишечника головастика при трансплантации в эну- клеированную зиготу могут вызывать развитие нормального зародыша. Эти результаты требовали объяснения. В настоящее время можно дать, по-видимому, два объяснения результатам Гердона. Первое: среди эпителиоцитов кишечника есть вдетки, которые могут подвергнуться дедифференцировке, стать снова полипотент- ными и обеспечить развитие зародыша. Второе: в процессе развития не все клетки дифференцируются, очень небольшая часть из них сохраняет экви- и полипотентность, составляя камбий различных тканей. Это можно объяснить небольшим процентом успешных трансплантаций у Гердона. Тем самым опыты Гердона не отменили основных выводов, сделанных Бриггсом и Кингом, но поставили вопрос о возможности дедифференцировки ядер в процессе эмбрионального развития.
Общий вывод, который можно сделать на основании всех Зтих экспериментов, состоит в том, что изменения в ядрах клеток имеют большое значение в механизме эмбриогенеза.
Возникает вопрос: какова роль цитоплазмы при этом? Для выяснения этого вопроса обратимся к результатам некоторых экспериментов. Хорошо известно, что образование всех видов РНК>
исключая митохоидриальные, протекает в ядрах клеток. Этот процесс транскрипции может быть легко обнаружен современными методами авторадиографии и биохимии. Как было показано выше,, становление синтеза различного вида молекул РНК происходит постепенно, по мере прохождения по стадиям развития. Этот процесс протекает только в ядре клеток.
Для решения вопроса в какой степени синтез молекул РНК зависит от зрелости цитоплазмы были поставлены следующие опыты. В энуклеированную зиготу трансплантировали ядра клеток,, выделенные у зародыша головастиков, находящихся на стадии бластулы, гаструлы и нейрулы. В этих ядрах, как известно, уже имеет место синтез различных комплексов молекул РНК. Однако, при взаимодействии этих ядер клеток с цитоплазмой зиготы они временно прекращали процессы транскрипции. Следовательно, реализации транскрипций на этапах эмбриогенеза б клетках зародыша требует определенной зрелости цитоплазмы, способной ингибиро- вать эти процессы.
Все сказанное выше позволяет сделать вывод по этому разделу: не только ядро определяет развитие элементарных актов в цитоплазме зародыша, но и цитоплазма в свою очередь регулирует ход элементарных актов в ядре клеток, т. е. взаимодействием ядра и цитоплазмы можно объяснить сложные процессы развития клеток, их дифференцировку на этапах эмбриогенеза.
Молекулярно-генетические механизмы дифференцировки клеток в последнее время подвергаются интенсивному изучению. У прокариот объяснение развития дифференциальной активности генов можно получить с позиций гипотезы Жакоба и Моно. Сложнее дело обстоит у эукариот. У бактерий, имеющих одну кольцевую хромосому, репрессия и депрессия генов получает объяснение с позиций взаимодействия оперона с геном-регулятором. У эукариотов в ядре находится несколько хромосом, в которых располагается громадная по объему генетическая информация. Существуют механизмы взаимодействия между отдельными порциями генетической информации, локализованными в различных хромосомах.
Приблизиться к пониманию хотя бы в обшюс чертах к механизмам взаимодействия между различными отделами генома клетки в процессе дифференцировок клеток эукариот при эмбриогенезе, помогает схема-гипотеза, высказанная Бриттеном^ Дэвидсоном.
В соответствии с понятиями Жакоба и Моно, Бриттен и Дэвидсон подразделяют гены на регуляторные и продюссерные (структурные). В свою очередь, регуляторные гены подразделяют на сензорные, интегрирующие и рецепторные. Сензорные гены взаимодействуют с регуляторными факторами-сигналами (гормоны,, биологически активные вещества). Сензорным генам подчинены гены-интеграторы, которые под воздействием сензорных генов определяют выработку активаторной РНК. Последняя может дейст
вовать на рецепторные гены, расположенные в различных «ролф- сомах и тем самым генерировать и усиливать сигнал. Рецепторные гены под действием активированной РНК включают гены- продюссеры, которые подобно структурным генам определяют синтез различных видов РНК, а, следовательно, и синтез определенных видов белка, которыми на конечном этапе и определяется ход и направление дифференцировки клеток.
Характер этих взаимодействий молекулярно-генетических механизмов клеток при их дифференцировке представлен на схеме 1.
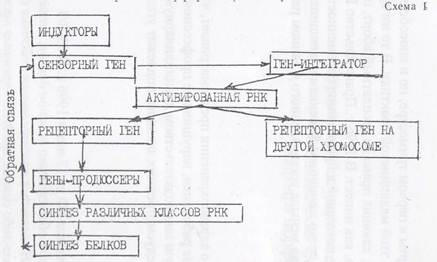
Некоторые из вновь синтезированных белков, или продукты,, возникающие под их действием, обеспечивают обратную связь,, воздействуя на сензорные гены.
Процесс развития протекает постепенно по схеме причинно- следственных взаимоотношений, следствие причины 1 может выступать в роли причины 2 и определять появление следствия 2 и т. д. Этот процесс развертывается во времени. Время, в свою очередь, определяется последовательностью генетически детерминированных процессов, протекающих в какой-то структуре.
Процесс развития зародышей не автономный, а зависит от действия ряда причин. Неоплодотворенные половые клетки у высших, организмов не способны обеспечить развитие зародышей. Толчком или причиной, обуславливающей развитие, является оплодотворение, которым запускается весь эмбриогенез и, в первую очередь,, дробление.
Развитие элементарных молекулярных актов на этапах раннего эмбриогенеза объясняется взаимодействием генетических структур ядра и органелл цитоплазмы. На стадии нейрулы основные
эмбриональные зачатки уже детерминированы, чем и подготовлен следующий этап в развитии зародыша—этап гистогенеза, а затем и органогенеза.
В развитии этих этапов важнейшая роль принадлежит взаимодействию клеток. Характер этих взаимодействий для высших животных был вскрыт в опытах Шлепана и его учеников. При этом было показано, что удаление зачатка хорды у зародышей амфибий блокирует образование из эктодермы нервной пластинки и превращение последней в нервную трубку. Пересадка хордального тяжа на вентральную сторону зародыша приводила к развитию нервной трубки на передней стороне тела головастика. Если зародышу головастика на стадии гаструлы трансплантировать зачаток хорды, взятый из другого зародыша, то отмечалось развитие двух нервных трубок на дорзальной и вентральной сторонах тела. На основании этих опытов был сделан вывод, что хорда вызывает развитие нервной трубки.
Этот процесс получил название эмбриональной индукции, а метаболиты хорды, ответственные за процесс, получили название индукторов.
Вторым классическим примером взаимодействий в процессах эмбрионального органогенеза может быть индукция развития хрусталика под действием глазного бокала. Индуктор является метаболитом и переносится из клеток индуцирующей в клетки индуцируемой структуры. Разделение этих структур с помощью сплошных, непроницаемых перегородок блокирует эффект индуктора, разделение с помощью решетчатых перегородок не препятствует развитию эффекта. Индуктор, достигнув клеток-мишеней, выступает в роли фактора, действующего на генетические структуры— сензорные гены и вызывает стойкое и наследуемое изменение активности генов — развитие эпигеномной наследственности. Таков, по-видимому, механизм детерминации, определяющий в дальнейшем характер последующих дифференцировок.
Коррекция процессов дифференцировки и перемещение клеток, приводящих к развитию тканевых и органных структур, получают объяснение с позиций принципа проверки обстановки клетками, предложенного Боннером. Согласно этому принципу, дифференци- ровка клеток идет ступенчато и клетки проверяют окружающую обстановку, сверяя ее с генетической программой своего развития. Если такая сверка совпадает с программной, то наступает следующий запрограммированный этап развития. Реализация принципа проверки обстановки допустима в том случае, если считать, что принцип дополняется принципом сигнализации. Согласно этому принципу каждый этап дифференцировки сопровождается выработкой клетками веществ сигналов, которые являются индукторами для последующих дифференцировок в клетках, проверяющих обстановку.
|
|
|
|
|
Дата добавления: 2014-12-17; Просмотров: 675; Нарушение авторских прав?; Мы поможем в написании вашей работы!