КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Влияние температуры на рост микроорганизмов
|
|
|
|
Температурные условия в биосфере достаточно разнообразны. Свыше 80% ее принадлежит к постоянно холодным областям. Значительная часть поверхности суши, включающая и континент Антарктиду, имеет низкую температуру. Средняя температура почвы в умеренной климатической зоне составляет 12°. Примерно 75% поверхности Земли приходится на долю Мирового океана, и около 90% его объема имеет температуру ниже 5°. Таков общий температурный профиль Земли.
Но на Земле есть много мест, резко различающихся по температурному режиму. Это области, где температура постоянно низкая (подземные и обледенелые пещеры, глубинные слои океанов) или высокая (действующие вулканы, выходы на поверхность земли струй паров и газов из расщелин или отверстий, кипящие или некипящие горячие источники, отходы различных технологических процессов). Есть также много областей с меняющимся температурным режимом: поверхностные слои морей и океанов, мелкие пресные водоемы и реки, верхние слои атмосферы, большинство мест на суше в зонах с умеренным и холодным климатом. Во многих областях с умеренным климатом температура колеблется от нуля и ниже до 30° и выше. В условиях холодного климата температурные колебания могут быть и более значительными.
Температура водных микроорганизмов равна температуре окружающей среды, поэтому воздействие температуры на клетки неизбежно. Температура влияет на скорость биохимических реакций, метаболические пути и состав биомассы.
При определении влияния температуры на прокариотные организмы следует различать два момента: способность организмов к выживанию после длительного нахождения в экстремальных температурных условиях и способность их к росту в этих условиях. Приспособления, сформированные у прокариот для перенесения неблагоприятных условий, в том числе и температурных, — это споры, цисты. Устойчивость вегетативных клеток и различных покоящихся форм больше в условиях воздействия низкими температурами. Так, вегетативные клетки и покоящиеся формы сохраняли жизнеспособность после длительного выдерживания при температуре, близкой к абсолютному нулю. Последнее используется в качестве одного из способов, обеспечивающих длительное хранение культур прокариот.
При изучении влияния температуры на рост прокариотных организмов выделяют температурный диапазон, ограниченный минимальной и максимальной температурами, при которых рост прекращается, а также область оптимальных температур с максимальной скоростью роста. Положение на температурной шкале основных точек (минимальная, максимальная, оптимальная температуры), а также величина температурного диапазона роста прокариот сильно различаются. На основании этих показателей прокариоты делят на три основные группы (см. табл. 2.2): психрофилы («любят» низкие температуры), мезофилы («любят» средние температуры) и термофилы (любят высокие температуры). Соответствующую терминологию используют и для обозначения температурного режима роста клеток, например «психрофильные условия», «термофильный режим», «биореактор мезофильного сбраживания» и т.д.
Большинство известных бактерий относится к мезофилам, которых обычно культивируют при температуре +37°С (середина оптимального диапазона температур), а температурный диапазон, в котором возможен рост, находится между 10 и 45-50°. Типичным мезофилом является Е. соli: нижняя граница роста +10°, верхняя +49°, оптимальная температура +37°.
Таблица 2.2
Классификация бактерий по отношению к температуре
| Наименование группы бактерий | Подгруппа | Температурный интервал роста (от … до …), °С | Диапазон оптимальных температур, °С |
| Психрофилы | Облигатные психрофилы | от –(10¸20) до +20 | 5¸15 |
| Факультативные психрофилы | от –10 до +(32¸35) | 20¸30 | |
| Мезофилы | - | от +10 до +(42¸45) | 30¸40 (чаще 35-38) |
| Факультативные термофилы | от +20 до +(70¸80) | 60¸70 | |
| Термофилы | Облигатные термофилы | от +(42¸42) до +(65¸70) | 50¸60 |
Психрофилы и факторы, определяющие возможность роста при низких температурах. Область температур роста психрофилов лежит в пределах от -10 до +20° и выше. Если при высоких температурах клетки погибают, низкие температуры (даже близкие к абсолютному нулю) они способны переносить длительное время (естественно, без роста и размножения). Это свойство микроорганизмов широко используют для их хранения. В области вечной мерзлоты были найдены бактерии с возрастом около миллиона лет, сохранившие способность возобновлять рост при переносе в благоприятные температурные условия.
Психрофилы делятся на облигатных и факультативных. Основное различие между подгруппами заключается в том, что облигатные психрофилы не способны к росту при температуре выше 20°, а верхняя температурная граница роста факультативных форм намного выше. Таким образом, факультативные психрофилы характеризуются более широким температурным диапазоном, при котором возможен их рост. И если в области низких температур они сходны с облигатными формами, то в области повышенных температур обладают способностью размножаться в значительно более высоких температурных границах. Различаются они также и оптимальными температурными зонами роста, находящимися у облигатных психрофилов значительно ниже, чем у факультативных (табл. 2.2). Принципиальное же сходство между ними — способность к росту при 0° и минусовых температурах.
Существование двух типов психрофилов объясняется особенностями их мест обитания. Облигатные психрофилы приспособились к устойчивым холодным условиям (глубины морей и океанов, ледяные пещеры). Напротив, психрофилы второго типа приспособились к обитанию в неустойчивых холодных условиях. В природе большинство психрофилов представлено факультативными формами. Способность психрофилов расти в условиях низких температур связывают в первую очередь с особенностями их ферментных белков и мембранных липидов. Увеличение в последних содержания ненасыщенных жирных кислот позволяет мембранам находиться в функционально активном жидкостно-кристаллическом состоянии при низких температурах. Обязательное условие возможности роста психрофилов при минусовых температурах — нахождение воды в жидком состоянии.
Термофилы и механизмы термофилии. Группу термофилов делят на 4 подгруппы:
1. Термотолерантные виды растут в пределах от 10 до 55 — 60°, оптимальная область лежит при 35 — 40°. Основное их отличие от мезофилов — способность расти при повышенных температурах, хотя.оптимальные температуры роста для обеих групп находятся на одном уровне.
2. Факультативные термофилы имеют максимальную температуру роста между 50 и 65°, но способны также к размножению при комнатной температуре (20°); оптимум приходится на область температур, близких к верхней границе роста. Особенность этой группы прокариот — способность к росту в области от 20 до 40°.
3. К облигатным термофилам относят виды, обнаруживающие способность расти при температурах около 70° и не растущие ниже 40°. Оптимальная температурная область облигатных термофилов примыкает к их верхней температурной границе роста. Представители этой подгруппы: эубактерии Bacillus acidocaldarius, Synechococcus lividus, архебактерии Methanobacterium thermoautotrophicum, Thermoplasma acidophilum и др.
4. Наконец, недавно обнаружены прокариоты, выделенные в подгруппу экстремальных термофилов и способные расти при температурах до 80-90°С с оптимумом в области 70-75°С. При высоком давлении, когда температура кипения воды превышает 100°С, рост возможен и при более высоких температурах. Например, в океане на глубине около 2,5 км в зоне горячего источника были обнаружены бактерии, способные расти при температуре 200-300°С и давлении 255 атм (при таком давлении вода закипает только при температуре 460°С). К экстремальным термофилам относятся организмы из группы архебактерии, не имеющие аналогов среди мезофилов, например представители родов Thermoproteus, Pyrococcus, Pyrodictium и др.
Разнообразие прокариот, которые удается культивировать при высоких или относительно высоких температурах, достаточно велико. Способность расти при температурах от 50 до 70°, свойственная представителям термотолерантных, факультативных и облигатных термофилов, не связана с осуществлением ими какого-либо одного специфического типа метаболизма. Среди термофилов, относящихся к этим подгруппам, найдены фотосинтезирующие, хемолитотрофные и хемогетеротрофные бактерии. Есть среди них облигатные аэробы и анаэробы. Термофилы, верхний предел роста которых ограничен 70°, в целом структурно напоминают своих мезофильных аналогов и по типам осуществляемого ими конструктивного и энергетического метаболизма относятся к тем же группам, что и мезофильные виды. По мере повышения температуры число видов, способных к росту, быстро уменьшается. Температурный предел для фотосинтезирующих эубактерии ограничен 70 — 73°. Это связывают с их неспособностью формировать функционально активные фотосинтетические мембраны.
Температурная ниша выше 70°, занятая экстремальными термофилами, гораздо беднее представителями. Верхний температурный предел, при котором зафиксирован рост в виде чистой бактериальной культуры в лаборатории, составляет 110°. Он обнаружен у архебактерии Pyrodictium occultum, растущей в диапазоне от 82 до 110° с оптимумом при 105°. Имеются также сообщения о том, что в природных условиях представители прокариот способны к росту при значительно более высоких температурах25.
Появились публикации об обнаружении бактерий, способных расти при температуре воды 250-300° и давлении 265 атм (при этом давлении Вода в жидком состоянии может находиться до 460°). Эти бактерии выделены из проб воды, поднятых с глубины 2560 м над поверхностью Тихого океана, где предположительно они существуют в горячих струях, выбрасываемых на дне океана так называемыми "черными гейзерами". Давление а районе обнаружения бактерий около 250 атм, а температура воды может быть выше 350°. В связи с этим исследователи начинают переоценивать Границы условий, при которых способны развиваться прокариоты. Высказывается предположение, что прокариоты могут существовать везде, где есть вода в жидком состоянии и достаточное количество питательных веществ.
Экстремальные термофилы относятся исключительно к архебактериям и представлены метанобразующими формами и видами, метаболизм которых связан с молекулярной серой. Почти все они — строгие анаэробы, но есть среди них и аэробы (представители рода Sulfolobus). Конструктивный метаболизм авто- или гетеротрофного типа. Анаэробные автотрофы получают энергию в результате восстановления CO2 или S молекулярным водородом с образованием в качестве конечных продуктов метана или сероводорода соответственно. Гетеротрофные экстремально термофильные анаэробы используют различные органические субстраты (белки, углеводы) для получения энергии в процессах брожения или анаэробного дыхания с молекулярной серой в качестве конечного акцептора электронов. Аэробные формы получают энергию в процессах, связанных с окислением молекулярной серы, железа или органических соединений. Большинство экстремальных термофилов не связано близким родством с нетермофильными изолятами.
Выяснение механизмов, обеспечивающих активное существование при высоких температурах, препятствующих в норме росту подавляющего большинства прокариот, представляет несомненный интерес. Проведенные в этом направлении исследования привели к убеждению, что термофилия включает множество молекулярных механизмов и не может быть объяснена только каким-нибудь одним свойством организма. Предложено несколько гипотез для объяснения природы термофилии. Одна из них подчеркивает роль мембранных липидов. Известно, что насыщенные жирные кислоты, входящие в состав липидов, имеют более высокую точку плавления по сравнению с ненасыщенными. Уже давно было замечено, что липиды термофилов имеют более высокие температуры плавления, чем липиды мезофилов, что достигается возрастанием содержания насыщенных жирных кислот в мембранах при повышении температуры культивирования. На основании этих данных и было высказано предположение, что липиды играют определенную роль в молекулярных механизмах термофилии, способствуя термостабильности мембран, и что нижняя температурная граница роста термофилов определяется температурой плавления мембранных липидов.
Многие исследователи считают, что определяющая роль в термофилии принадлежит белкам, в первую очередь ферментным. С этих позиций основные температурные точки термофилов зависят от конформации одного или нескольких ключевых ферментов: при минимальной температуре роста происходит переход от жесткой неактивной конформации белковых молекул к конформации с ограниченной гибкостью; оптимальная температура роста определяет наиболее благоприятное конформационное состояние ферментных белков; при максимальной температуре начинаются нарушения конформации белков и снижение их ферментативной активности, а выше этой температуры рост прекращается вследствие тепловой денатурации белков.
В одной из гипотез термофилии постулируется термостабильность структурных компонентов клетки термофилов. Оказалось, что клеточная стенка, мембраны, рибосомы термофилов, значительно более термостабильны, чем соответствующие структуры мезофилов. Особенно большое внимание в этом плане привлекают клеточные мембраны.
Разобранные выше гипотезы, вероятно, вносят определенный вклад в механизм термофилии, дополняя друг друга, хотя не исключено, что молекулярные механизмы этого явления могут быть значительно шире и для их понимания необходимы дополнительные исследования. В природе нет четкого деления на группы, представленные в табл. 2.2, т.к. оптимальные температурные зоны для разных групп образуют почти непрерывный ряд температур, а температурные диапазоны, допускающие рост выделенных групп, значительно перекрываются.
Удельная скорость роста определяется скоростью биохимических реакций 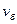 , которую можно выразить как произведение скорости химической реакции
, которую можно выразить как произведение скорости химической реакции 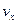 и активности ферментов
и активности ферментов  , представленной как коэффициент ускорения реакции:
, представленной как коэффициент ускорения реакции:
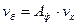 (2.42)
(2.42)
Скорость химической реакции возрастает с повышением температуры согласно уравнению Аррениуса. Активность ферментов проявляется только в определенном интервале температур, внутри которого она возрастает, достигая максимума, а затем падает. В результате, зависимость скорости биохимических реакций и удельной скорости роста клеток от температуры имеет вид несимметричной кривой с максимумом (рис. 2.8).
Активность ферментов постоянна (максимальна) в интервале температур ТА. Согласно уравнению (2.42), в этом температурном интервале удельная скорость роста (как интегральная характеристика скорости биохимических реакций) зависит от температуры по уравнению Аррениуса:
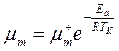 (2.43)
(2.43)
где 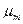 - удельная скорость роста при отсутствии лимитирования роста (максимальная удельная скорость роста);
- удельная скорость роста при отсутствии лимитирования роста (максимальная удельная скорость роста); 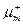 – константа уравнения;
– константа уравнения;  – энергия активации, кДж/моль (ккал/моль);
– энергия активации, кДж/моль (ккал/моль); 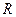 – универсальная газовая постоянная (8,31×10-3 кДж/моль×K или
– универсальная газовая постоянная (8,31×10-3 кДж/моль×K или
1,98×10-3 ккал/моль×K); 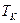 – абсолютная температура, K).
– абсолютная температура, K).

Рис. 2.8. Зависимость удельной скорости роста от температуры при отсутствии лимитирования роста (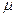 = ):
= ):
Троста – температурный интервал роста;
Tопт – интервал оптимальных температур, где достигается наибольшая скорость роста;
ТА – интервал температур, где выполняется уравнение Аррениуса.
Для бактерий величина энергии активации составляет 50-70 кДж/моль (12-17 ккал/моль), соответственно скорость роста примерно удваивается при увеличении температуры на 10°. Температурный интервал ТА, где справедливо уравнение Аррениуса, обычно не превышает 10-15°С. В более широком интервале температур используют эмпирические зависимости:
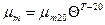 , (2.44)
, (2.44)
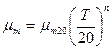 , (2.45)
, (2.45)
где 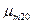 – максимальная удельная скорость роста при температуре 20°С;
– максимальная удельная скорость роста при температуре 20°С; 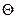 ,
, 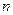 - эмпирические константы ( =1,04-1,12; =1-2,5);
- эмпирические константы ( =1,04-1,12; =1-2,5); 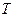 - температура, °С.
- температура, °С.
С увеличением температуры одновременно с ростом происходит увеличение константы Моно 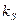 . Поэтому при низких концентрациях субстрата (
. Поэтому при низких концентрациях субстрата ( < , резкое лимитирование роста по субстрату) возможно падение удельной скорости роста с повышением температуры даже в интервале, где справедливо уравнение Аррениуса. Действительно, при < выполняется:
< , резкое лимитирование роста по субстрату) возможно падение удельной скорости роста с повышением температуры даже в интервале, где справедливо уравнение Аррениуса. Действительно, при < выполняется:
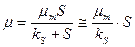 . (2.46)
. (2.46)
Если при увеличении температуры константа возрастает в большей степени, чем , то отношение 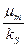 будет падать, соответственно будет уменьшаться удельная скорость роста . Отсюда следует, что характер зависимости
будет падать, соответственно будет уменьшаться удельная скорость роста . Отсюда следует, что характер зависимости 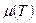 может отличаться от зависимости
может отличаться от зависимости 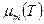 , что иллюстрирует рис. 2.9.
, что иллюстрирует рис. 2.9.
С повышением температуры растут затраты на поддержание жизнедеятельности (увеличивается коэффициент энергии поддержания 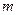 и константа скорости самоокисления
и константа скорости самоокисления 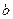 ). При лимитировании роста по субстрату увеличение с повышением температуры может быть незначительным (возможно даже падение - см. рис. 2.9), поэтому возрастание коэффициентов и вызовет уменьшение экономического коэффициента (
). При лимитировании роста по субстрату увеличение с повышением температуры может быть незначительным (возможно даже падение - см. рис. 2.9), поэтому возрастание коэффициентов и вызовет уменьшение экономического коэффициента (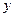 ) и увеличение энергетического коэффициента (
) и увеличение энергетического коэффициента (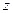 ) в соответствие с уравнениями (2.26), (3.32) и (3.35), (3.36). Данная закономерность проявляется в сооружениях биологической очистки воды, где летом (при более высоких температурах) прирост биомассы ниже, чем зимой.
) в соответствие с уравнениями (2.26), (3.32) и (3.35), (3.36). Данная закономерность проявляется в сооружениях биологической очистки воды, где летом (при более высоких температурах) прирост биомассы ниже, чем зимой.

Рис. 2.9. Зависимость удельной скорости роста от температуры при лимитировании роста по субстрату:
1 – растет с увеличением ; 2 – падает с увеличением .
Если рост клеток лимитирован по диффузии субстрата (или кислорода), то влияние температуры на скорость роста существенно изменяется, ибо оно отвечает зависимости скорости диффузии от температуры. Скорость диффузии возрастает с увеличением температуры в соответствии с увеличением коэффициента диффузии. На практике можно использовать уравнения:
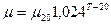 , (2.47)
, (2.47)
 . (2.48)
. (2.48)
Если для описания зависимости при лимитировании скорости роста по диффузии применить уравнение Аррениуса, то величина энергии активации обычно не превышает 6 ккал/моль. Это существенно ниже, чем в случае отсутствия лимитирования роста диффузией, что позволяет по температурной зависимости выявлять фактор, лимитирующий рост. Обычно полагают, что если найденное экспериментально значение < 8 ккал/моль, то рост лимитирован по диффузии. Если > 10 ккал/моль, то лимитирования роста диффузией нет.
|
|
|
|
|
Дата добавления: 2014-12-27; Просмотров: 8481; Нарушение авторских прав?; Мы поможем в написании вашей работы!