КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Теория хемостата
|
|
|
|
Классификация методов культивирования
КУЛЬТИВИРОВАНИЕ КЛЕТОК
Влияние азота и фосфора на рост микроорганизмов
Азот и фосфор относятся к важнейшим биогенным элементам, необходимым для роста всех клеток. Азот необходим для конструктивного обмена (синтеза белков), фосфор используется в конструктивном и энергетическом обменах, входя в состав таких биомолекул, как ДНК, РНК, фосфолипиды и АТФ.
Зависимость удельной скорости роста от концентрации азота и фосфора подчиняется уравнению Моно при величине константы Моно по азоту 0,1-0,5 мг/л, по фосфору 0,1-0,2 мг/л.
При лимитировании роста по азоту, а также одновременно по азоту и фосфору, клетки часто переходят на синтез безазотистых соединений, т.е. вместо белков синтезируются полисахариды, липиды и поли-b-оксимасляная кислота. Количество безазотистых соединений в клетке становится значительно больше нормального уровня, поэтому говорят о сверхсинтезе безазотистых соединений.
Данное явление используется в биотехнологии, например, для получения полисахаридов. В системах биологической очистки воды сверхсинтез полисахаридов играет отрицательную роль, ухудшая способность бактериальных хлопьев (хлопьев активного ила) к осаждению.
В условиях сверхсинтеза безазотистых соединений экономический коэффициент достигает аномально высоких значений, превосходящих величину истинного экономического коэффициента. Это связано с низкими затратами энергии на биосинтез полисахаридов и липидов, чем на биосинтез белков, соответственно, меньшая часть субстрата вовлекается в энергетический обмен (окисляется для получения энергии), и основная часть субстрата трансформируется в биомассу в ходе конструктивного обмена. Но образующаяся биомасса имеет аномальный состав с преобладанием тех или иных безазотистых соединений. Так, содержание липидов может достигать 70 % против 5-10 % в норме, а полисахаридов – до 80 %, против обычного их содержания в биомассе клеток около 15 %.
В зависимости от режима процесса выделяют периодическое и непрерывное культивирование.
Периодическое культивирование подразделяют на простое и отъемно-доливное. При простом периодическом культивировании в биореактор загружают свежую питательную среду и клетки, проводят процесс определенное время без добавок субстрата и отвода культуральной жидкости, затем реактор опорожняют и вновь повторяют цикл культивирования. Отъемно-доливное культивирование отличается тем, что после завершения процесса реактор полностью не опорожняют, а лишь удаляют (отнимают) часть культуральной жидкости (клетки остаются в реакторе) и затем доливают в реактор такой же объем свежей питательной среды.
Периодическое культивирование легко реализовать как в лабораторных, так и в промышленных условиях, но на практике оно применяется все же реже, так уступает непрерывному культивированию по производительности и стабильности. Из систем биологической очистки в режиме отъемно-доливного культивирования работают аэротенки периодического действия, иногда применяемые для очистки небольших объемов сточных вод.
Непрерывное (проточное) культивирование – это метод, при котором в биореактор непрерывно подается раствор субстрата и с тем же объемным расходом отводится суспензия клеток или культуральная жидкость. В зависимости от гидродинамического режима в биореакторе выделяют два основных типа непрерывного культивирования: тубулярная культура и хемостат. В тубулярной культуре биореактор по гидродинамическому режиму является идеальным вытеснителем. Хемостат – это установка непрерывного культивирования в реакторе идеального смешения при постоянной скорости разбавления.
В проточном реакторе-вытеснителе (тубулярной культуре) продольное перемешивание практически отсутствует, при этом условия среды и состояние микроорганизмов изменяются по длине реактора так же, как с течением времени в периодической культуре (рис. 3.1 а). На этой основе в лабораторных условиях тубулярную культуру часто моделируют путем периодического культивирования. Например, аэротенк-вытеснитель моделируется отъемно-доливным культивированием.
| а) | 
|
| б) | 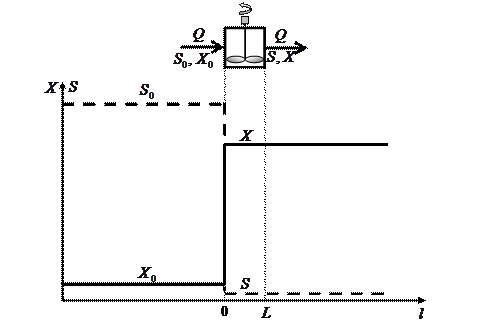
|
| в) | 
|
| г) | 
|
| Условные обозначения: | 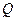 - расход; - расход;  - концентрация субстрата; - концентрация субстрата; 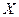 - концентрация биомассы; - концентрация биомассы; 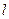 - линейная координата; - линейная координата; 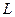 - длина биореактора. - длина биореактора.
|
Рис. 3.1. Схемы непрерывного культивирования: а) тубулярная культура без возврата биомассы; б) хемостат без возврата биомассы; в) батарея хемостатов; г) непрерывное культивирование с возвратом биомассы (система биологической очистки аэротенк-отстойник).
Принципиально другими свойствами обладает хемостат, позволяющий в течение длительного времени поддерживать стабильные условия среды и стабильное физиологическое состояние микроорганизмов (рис. 3.1 б). Кроме того, хемостат может устойчиво работать даже при отстутствии микроорганизмов в поступающей питательной среде.
Последовательно соединенные реакторы идеального смешения (батарея хемостатов) образует систему, приближающуюся по гидродинамическому режиму к идеальному вытеснению (рис. 3.1 в). Поэтому на практике биореактор-вытеснитель часто представляет собой батарею хемостатов.
Непрерывное культивирование может осуществляться без возврата и с возвратом биомассы в реактор. В режиме непрерывного культивирования без возврата биомассы работают биопруды, применяемые для доочистки сточных вод. Аэротенки относятся к реакторам непрерывного культивирования с возвратом биомассы. Микроорганизмы в аэротенках находятся в виде агрегатов, состоящих из тысяч слипшихся клеток. Эти агрегаты, называемые хлопьями активного ила, хорошо оседают при отстаивании. Поэтому для отделения биомассы от воды обычно используют отстойники. Аэротенк вместе с отстойником образует единую систему биологической очистки, работающую в режиме непрерывного культивирования с возвратом биомассы (рис. 3.1 г).
Клетки в биореакторах могут находиться в различных формах: суспендированные дисперсные клетки, суспендированные клеточные агрегаты (например, хлопья активного ила), иммобилизованные клетки (например, биопленки на твердых поверхностях), биогранулы размером 1-2 мм, состоящие из множества клеток.
В зависимости от кислородного режима выделяют аэробное и анаэробное культивирование. Кроме того, при биологической очистке от азота используют аноксидное культивирование (в биореакторе отсутствует растворенный кислород, но имеется химически связанный кислород в виде нитритов и нитратов).
Хемостат без возврата биомассы. Изменение количества биомассы в хемостате определяется ее приростом, поступлением с потоком питательной среды и оттоком из реактора. Для бесконечно малого промежутка времени 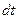 этот баланс задается уравнением:
этот баланс задается уравнением:
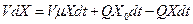 , (3.1)
, (3.1)
где 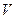 - гидравлический объем хемостата, м3; - концентрация биомассы в хемостате, кг/м3;
- гидравлический объем хемостата, м3; - концентрация биомассы в хемостате, кг/м3; 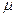 - удельная скорость роста клеток, ч-1; - объемный расход питательной среды, м3/ч;
- удельная скорость роста клеток, ч-1; - объемный расход питательной среды, м3/ч; 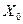 - концентрация биомассы в питательной среде, подаваемой в хемостат, кг/м3.
- концентрация биомассы в питательной среде, подаваемой в хемостат, кг/м3.
Поделив на 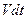 , получим:
, получим:
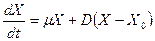 , (3.2)
, (3.2)
где 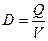 - скорость разбавления, ч-1.
- скорость разбавления, ч-1.
При =0
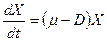 . (3.3)
. (3.3)
Отсюда вытекает важное свойство хемостата – автоматическая стабилизация режима. Действительно, если повысится сверх заданной нами величины 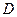 , то согласно (3.3), при
, то согласно (3.3), при 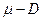 >0, скорость изменения концентрации биомассы в хемостате
>0, скорость изменения концентрации биомассы в хемостате 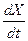 >0. Следовательно, величина будет возрастать, что повлечет снижение , которая по определению обратно пропорциональна (зависимость (2.1)). Аналогичным образом, если окажется < , то это вызовет увеличение .
>0. Следовательно, величина будет возрастать, что повлечет снижение , которая по определению обратно пропорциональна (зависимость (2.1)). Аналогичным образом, если окажется < , то это вызовет увеличение .
Таким образом, при отклонении от задаваемой скорости разбавления , хемостат автоматически возвращает значение , поддерживая равенство = , отвечающее стационарному режиму.
Баланс по субстрату и продукту приводит к уравнениям:
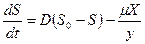 , (3.4)
, (3.4)
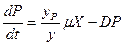 , (3.5)
, (3.5)
где  , - концентрация субстрата соответственно в поступающей питательной среде и в хемостате, кг/м3;
, - концентрация субстрата соответственно в поступающей питательной среде и в хемостате, кг/м3; 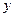 - экономический коэффициент (разд. 2.1);
- экономический коэффициент (разд. 2.1); 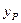 - выход продукта (разд. 2.1);
- выход продукта (разд. 2.1);  - концентрация продукта в хемостате, кг/м3.
- концентрация продукта в хемостате, кг/м3.
Как следует из (3.3)-(3.5), в стационарном режиме ( =0, 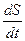 =0,
=0, 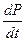 =0):
=0):
= , (3.6)
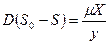 , (3.7)
, (3.7)
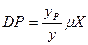 . (3.8)
. (3.8)
Если зависимость от подчиняется уравнению Моно (разд. 2.3), то подставляя 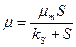 , получим выражения для стационарных концентраций биомассы, субстрата и продукта в хемостате:
, получим выражения для стационарных концентраций биомассы, субстрата и продукта в хемостате:
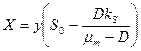 , (3.9)
, (3.9)
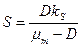 , (3.10)
, (3.10)
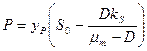 . (3.11)
. (3.11)
Как видно из этих уравнений, стационарные концентрации биомассы и продукта линейно возрастают с увеличением концентрации субстрата в поступающем потоке питательной среды . Напротив, стационарная концентрация субстрата в хемостате не зависит от величины и целиком определяется задаваемой нами скоростью разбавления и константой Моно 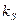 .
.
С увеличением скорости разбавления концентрация субстрата возрастает, стремясь к начальной величине , а концентрация биомассы и продукта падает до нуля. Скорость разбавления, при которой рост в хемостате прекращается (соответственно, = , =0, =0), называется критической. Величина критической скорости разбавления 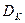 находится из уравнений (3.9)-(3.11). Например, из (3.9), при =0, получим:
находится из уравнений (3.9)-(3.11). Например, из (3.9), при =0, получим:
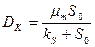 . (3.12)
. (3.12)
Обычно >> , поэтому в первом приближении выполняется: 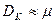 .
.
С приближением скорости разбавления к критической (в области 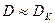 ) рост в хемостате невозможен. На этом основан отбор видов в хемостате. Принцип отбора видов состоит в следующем. Если в хемостат загрузить смешанную культуру, состоящую из нескольких видов микроорганизмов, то после включения хемостата при скорости разбавления в нем останутся только те виды, для которых выполняется: > . Остальные виды, для которых £ , покинут хемостат. Увеличивая величину , можно добиться того, что в хемостате останется только один, самый быстрорастущий вид микроорганизимов.
) рост в хемостате невозможен. На этом основан отбор видов в хемостате. Принцип отбора видов состоит в следующем. Если в хемостат загрузить смешанную культуру, состоящую из нескольких видов микроорганизмов, то после включения хемостата при скорости разбавления в нем останутся только те виды, для которых выполняется: > . Остальные виды, для которых £ , покинут хемостат. Увеличивая величину , можно добиться того, что в хемостате останется только один, самый быстрорастущий вид микроорганизимов.
При ингибировании роста уравнения (3.6)-(3.8) сохраняются, но зависимость 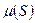 не подчиняется уравнению Моно. Соответственно, изменяются выражения для стационарных концентраций биомассы, субстрата и продукта в хемостате.
не подчиняется уравнению Моно. Соответственно, изменяются выражения для стационарных концентраций биомассы, субстрата и продукта в хемостате.
Хемостат с возвратом биомассы. Возврат или рециркуляция биомассы (рис. 3.1 г) увеличивает ее концентрацию в биореакторе, что повышает скорость потребления субстрата. В хемостате без возврата биомассы, как следует из уравнений (3.6) и (3.7):
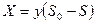 . (3.13)
. (3.13)
В хемостате с возвратом биомассы ее концентрация 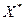 в биореакторе:
в биореакторе:
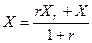 , (3.14)
, (3.14)
где 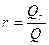 - коэффициент рециркуляции биомассы (рис. 4.1 г),
- коэффициент рециркуляции биомассы (рис. 4.1 г), 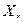 - концентрация возвращаемой биомассы, кг/м3.
- концентрация возвращаемой биомассы, кг/м3.
Для аэротенков-смесителей отношение 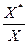 =20-50, т.е. возврат биомассы позволяет в десятки раз повысить концентрацию активного ила и, следовательно, скорость биологической очистки.
=20-50, т.е. возврат биомассы позволяет в десятки раз повысить концентрацию активного ила и, следовательно, скорость биологической очистки.
Системы биологической очистки работают в условиях нестабильности расхода поступающей воды и концентрации примесей, служащих субстратом для микроорганизмов. В этих условиях требуется значительно увеличить критическую скорость разбавления с тем, чтобы неравенство > надежно выполнялось во всем диапазоне изменений величины , зависящей от расхода воды. Это и достигается при введении возврата биомассы, который позволяет в десятки раз увеличить критическую скорость разбавления:
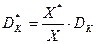 , (3.15)
, (3.15)
где 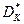 , - соответственно, критическая скорость разбавления в хемостате с возвратом биомассы и без возврата биомассы, ч-1.
, - соответственно, критическая скорость разбавления в хемостате с возвратом биомассы и без возврата биомассы, ч-1.
|
|
|
|
|
Дата добавления: 2014-12-27; Просмотров: 4676; Нарушение авторских прав?; Мы поможем в написании вашей работы!