КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Структура нейрона
|
|
|
|
Нейрон состоит из тела клетки, многочисленных ветвящихся коротких отростков — дендритов и одного длинного отростка — аксона, длина которого может достигать нескольких десятков сантиметров (рис. 18.1).
Объем цитоплазмы, содержащейся в отростках нервной клетки, может в несколько раз превышать ее количество в теле клетки. Тело нейрона окружено плазматической мембраной — плазмалеммой (рис. 18.2). В тесной связи с плазмалеммой1 в теле нейрона и проксимальных отрезках дендритов находится так называемая подповерхностная мембранная структура. Это — цистерны, которые расположены параллельно поверхности плазмалеммы и отделены от нее очень узкой светлой зоной. Предполагай?*!-, что цистерны играют важную роль в мета-
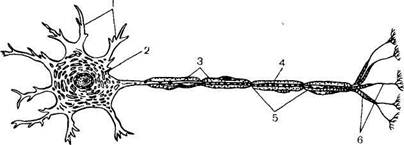
Рис. 18.1. Строение нейрона (схема по Шмитту).
1—дендриты; 2 - тело нейрона; 3—аксон; 4 — мделиновая оболочка; 5 — перехваты узла; 6 — окончания.
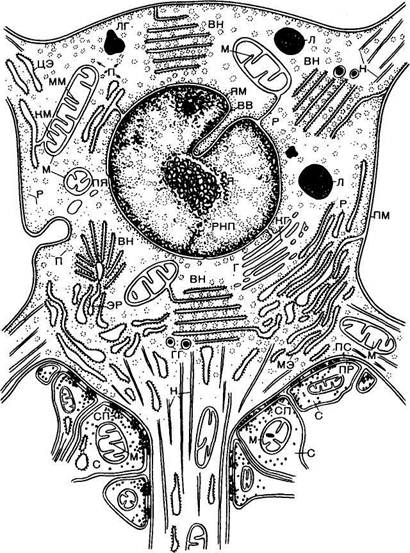
Рис. 18.2. Схематическое изображение ультратонкого строения нервной клетки по данным электронной микроскопии (по А. А. Маниной).
ВВ — впячивание ядерных мембран; ВН — вещество Ниссля; Г — пластинчатый комплекс (аппарат Гольд-жи); ГТ — гранулы гликогена; КГ — канальцы пластинчатого комплекса; КМ — кристы митохондрий; Л — лизосомы; ЛГ — липидные гранулы; М — митохондрии; ММ — мембрана митохондрий; МЭ — мембраны эндоплазматической сети; Н — нейропрофибриллы; П —полисомы; ПМ - плазматическая мембрана; ПР — пресинаптическая мембрана; ПС — постсинаптическая мембрана; ПЯ — поры ядерной мембраны; Р — рибосомы; РНП — рибонуклеопротеиновые гранулы; С — синапс; СП — синаптические пузырьки; ЦЭ — цистерны эндоплазматической сети; ЭР — эндоплазмагическая сеть; Я —ядро; ЯМ — ядерная мембрана.
болизме нейрона. Основной ультраструктурой цитоплазмы нейрона является эндоплазматиче-ская сеть — система ограниченных мембраной пузырьков, трубочек и уплощенных мешочков, или цистерн. Мембраны эндоплазматической сети связаны определенным образом с плазма-леммой и оболочкой ядра нейрона.
Гранулы, локализованные на мембранах эндоплазматической сети, а также свободно расположенные в цитоплазме, являются рибосомами.
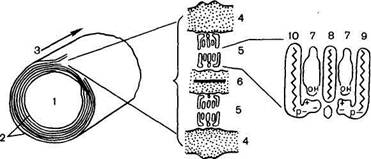
Рис. 18.3. Молекулярная организация миелиновой оболочки (по X. Хидену).
1 —аксон; 2 — миелин; 3 — ось волокна; 4 — белок (наружные слои); 5 —липиды; 6 — белок (внутренний слой); 7 — холестерин; 8 — цереброчид; 9 — сфингомиелин; 10 — фосфатидилсерин.
Характерной структурной основой нервной клетки является базофильное вещество (вещество, субстанция Ниссля), состоящее из рибонуклеиновых кислот и белков. В цитоплазме также выявляется сеть тонких нитей — нейрофибрилл, которые в совокупности образуют густую сеть. Нейрофибриллы — это структурное выражение правильной линейной ориентации белковых молекул.
Важный компонент цитоплазмы нейрона — пластинчатый комплекс (аппарат Гольджи), где сосредоточены главным образом липидные компоненты клетки. Одной из особенностей митохондрий, изолированных из нервных клеток, является то, что они содержат меньше ферментов, участвующих в процессах окисления жирных кислот и аминокислот, чем митохондрии из других тканей.
В 1ДНС лизосомы обнаруживаются постоянно и выполняют те же функции, что и лизо-сомы других органов и тканей.
Размер ядра нейрона колеблется от 3 до 18 мкм, достигая в крупных нейронах '/4 величины их тела.
Строение миелина
Нервные волокна, образующиеся из аксонов нервных клеток, по своему строению могут быть подразделены на два типа: миелиновые (мякотные) и безмиелиновые (бедные миелином). Проводниковая система соматической нервной системы, а также ЦНС относятся к первому типу, функционально более совершенному, обладающему способностью с высокой скоростью передавать нервные импульсы.
Миелиновое вещество — понятие чисто морфологическое. По сути миелин — система, образованная многократно наслаивающимися мембранами клеток нейроглии ' вокруг нервных отростков (в периферических нервных стволах нейроглия представлена леммоцитами, или шванновскими клетками, а в белом веществе ЦНС — астроцитами).
По химическому составу миелиновое вещество является сложным белково-липид-ным комплексом.
На долю липидов приходится до 80% плотного остатка; 90% всех липидов миелина представлены холестерином, фосфолипидами и цереброзидами. Считают, что в липоидных слоях миелиновых оболочек молекулы различных липидов имеют строго определенное расположение (рис. 18.3).
ХИМИЧЕСКИЙ СОСТАВ МОЗГА
Серое вещество головного мозга представлено в основном телами нейронов, а белое вещество — аксонами. В связи с этим указанные отделы мозга значительно отличаются по своему химическому составу. Эти отличия носят прежде всего количественный характер. Содержание воды в сером веществе головного мозга заметно больше, чем в белом веществе (табл. 18.1).
В сером веществе белки составляют половину плотных веществ, а в белом веществе — одну треть!. На долю липидов в белом веществе приходится более половины сухого остатка, в сером веществе — лишь около 30 %.
Таблица 18.1. Химический состав серого и белого вещества головного мозга человека (в процентах от массы сырой ткани)
| Составные части | Серое вещество | Белое вещество |
| Вода Сухой остаток Белки Липиды Минеральные вещества | 84 16 8 5 1 | 70 30 9 17 |
Белки
На долю белков приходится примерно 40% сухой массы головного мозга. Мозговая ткань является трудным объектом для изучения белкового состава вследствие большого содержания липидов и наличия белково-липидных комплексов.
Впервые А. Я. Данилевский разделил белки мозговой ткани на растворимые в воде и солевых растворах белки и нерастворимые белки. Обширные исследования в этой области были проведены также А. В. Палладиным и сотр., которые разделили белки нервной ткани на четыре фракции: извлекаемые водой; 4,5% раствором КС1; 0,1% раствором NaOH; нерастворимый остаток. Установлено, что серое вещество богаче белками, растворимыми в воде, чем белое вещество: соответственно 30 и 19%. Белое вещество, напротив, содержит гораздо больше (22%) нерастворимого белкового остатка, чем серое вещество (5%). В дальнейшем было выделено 5-10 фракций растворимых белков мозга, отличающихся по своей электрофорети-ческой подвижности.
В настоящее время, сочетая методы экстракции буферными растворами, хроматографии на колонках с ДЭАЭ-целлюлозой и диск-электрофореза в гюлиакриламидном геле, удалось из ткани мозга выделить около 100 различных растворимых белковых фракций.
В нервной ткани содержатся как простые, так и сложные белки. Простые белки — альбумины (нейроальбумины), глобулины (нейроглобулины), катионные белки (гис-тоны и др.) и опорные белки (нейросклеропротеины),
Поскольку альбумины и глобулины по своим физико-химическим свойствам несколько отличаются от аналогичных белков сыворотки крови, они, как правило, называются нейроальбуминами и нейроглобу.чинами. Количество нейроглобулинов в головном мозге относительно невелико — в среднем 5 % по отношению ко всем растворимым белкам. Нейроальбумины являются основным белковым компонентом фос-фопротеинов нервной ткани, на их долю приходится основная масса растворимых белков (89 — 90%). В свободном состоянии нейроальбумины встречаются редко. В частности, они легко соединяются с липидами, нуклеиновыми кислотами, углеводами и другими небелковыми компонентами.
Белки, которые в процессе электрофоре!ического разделения при рН 10,5-12,0 движутся к катоду, получили название катионных. Главнейшими представителями
этой группы белков в нервной ткани являются гистоны, которые делятся на пять основных фракций в зависимости от содержания в их полипептидных цепях остатков лизина, аргинина и глицина.
Ненросклеропротеины можно охарактеризовать как структурно-опорные белки. Основные представители этих белков — нейроколлагены, нейроэластины, нейростро-мины и др. Они составляют примерно 8 — 10% от всех простых белков нервной ткани и локализованы в основном в белом веществе головного мозга и в периферической нервной системе.
Сложные белки нервной ткани представлены нуклеопротеинами, липопротеинами, протеолипидами, фосфопрогеинами, гликопротеинами и т. д. В мозговой ткани содержатся в значительном количестве еще более сложные надмолекулярные образования, такие как липонуклеопротеины, липогликопротеины, возможно, и липоглико-нуклеопротеиновые комплексы.
Нуклеопротеины — белки, которые принадлежат либо к ДНП, либо к РНП. Часть этих белков из мозговой ткани извлекается водой, другая часть — солевыми средами, а третья —0,1 М раствором щелочи.
Липопротеины составляют значительную часть водорастворимых белков мозговой ткани. Их липидный компонент состоит в основном из фосфоглицеридов и холестерина.
Протеолипиды — единственные сложные белки, которые извлекаются органическими растворителями, например смесью хлороформа и метанола. В отличие от липопротеинов в них липидный компонент преобладает над белковым. Наибольшее количество протеолипидов сосредоточено в миелине, в небольших количествах они входят в состав синаптических мембран и синаптических пузырьков.
Фосфопротеины в головном мозге содержатся в большем количестве, чем в других органах и тканях, — около 2 % по отношению ко всем сложным белкам мозга. Фосфопротеины обнаружены в мембранах различных морфологических структур нервной ткани.
Гликогфотеины представляют собой чрезвычайно гетерогенную группу белков. По количеству белка и углеводов, входящих в состав гликопротеинов, их можно разделить на две основные группы. Первая группа — это гликопротеины, содержащие от 5 до 40% углеводов и их производных; белковая часть состоит преимущественно из альбуминов и глобулинов. В гликопротеинах, составляющих вторую группу, содержится 40—85% углеводов, часто обнаруживается липидный компонент; по своему составу они могут быть отнесены к гликолипопротеинам.
В последние годы в нервной ткани обнаружен ряд специфических белков. К таким белкам, в частности, относятся белок S-100 и белок 14-3-2. Белок S-100, или белок Мура, называют также кислым белком, так как он содержит большое количество остатков глутаминовой и аспарагиновой кислот. Этот белок сосредоточен в основном в нейроглии (85 — 90%), в нейронах его не более 10—15% от общего количества в головном мозге. Установлено, что концентрация белка S-100 возрастает при обучении (тренировках) животных. Однако пока нет оснований считать, что белок S-100 непосредственно участвует в формировании и хранении памяти. Не исключено, что его участие в этих процессах опосредованно. Белок 14-3-2 также относится к кислым белкам. В отличие от белка S-100 он локализован в основном в нейронах: в нейроглиальных клетках его содержание невелико. Пока неясна роль белка 14-3-2 в выполнении специфических функций нервной ткани.
Ферменты. В мозговой ткани содержится большое количество ферментов, катализирующих обмен углеводов, липидов и белков. Однако до сих пор в кристаллическом виде из ЦНС млекопитающих выделены лишь некоторые ферменты, в частности ацетилхолинэстераза и креатинкиназа.
Значительное количество ферментов в мозговой ткани находится в нескольких молекулярных формах (изоферменты): ЛДГ, альдолаза, креатинкиназа, гексокиназа, малатдегидрогеназа, глутаматдегидрогеназа, холинэстераза, кислая фосфатаза, моно-аминоксидаза и другие.
Липиды
Среди химических компонентов головного мозга особое место занимают липиды, высокое содержание и специфическая природа которых придают мозговой ткани характерные особенности. В группу липидов головного мозга входят фосфоглицериды, холестерин, сфингомиелины, цереброзиды, ган-глиозиды и очень небольшое количество нейтрального жира (табл. 18.2). Кроме того, многие липиды нервной ткани находятся в тесной взаимосвязи с белками, образуя сложные системы типа протеоли-пидов.
В сером веществе головного мозга фосфоглицериды составляют более 60 % от всех липидов, а в белом веществе - около 40 %. Напротив, в белом веществе содержание холестерина, сфингомиелинов и особенно переброзидов больше, чем в сером веществе.
| Холестерин | 22,0 | 27,5 | 27,7 |
| Цереброзиды | 5,4 | 19,8 | 22,7 |
| Ганглиозиды | 1,7 | 5,4 | 3,8 |
| Фосфа тидилэтаноламины | 22,7 | 14,9 | 15,6 |
| Фосфатидилхолины | 26,7 | 12,8 | 11,2 |
| Фосфатидилссрины | 8,7 | 7,9 | 4,8 |
| Фосфатидилинозитолы | 2 7 | 0,9 | 0,6 |
| Плазмалогены | 8^8 | 11,2 | 12,3 |
| Сфингомиелины | 6,9 | 7,7 | 7,9 |
Углеводы
В мозговой ткани имеются гликоген и глюкоза. Однако по сравнению с другими тканями гкань мозга бедна углеводами. Общее содержание глюкозы в головном мозге разных животных составляет в среднем 1—4 мкмоль на 1 г ткани, а гликоген - 2,5 - 4,5 мкмоль на 1 г ткани (в расчете на глюкозу). Интересно отметить, что общее содержание гликогена в мозге эмбрионов и новорожденных животных значительно выше, чем в мозге взрослых. Например, у новорожденных мышей в отличие от взрослых особей уровень гликогена в 3 раза выше. По мере роста и дифференцировки мозга концентрация гликогена быстро снижается и остается относительно постоянной у взрослого животного.
В мозговой ткани имеются также промежуточные продукты обмена углеводов: гексозо- и триозофосфаты, молочная, пировиноградная и другие кислоты. В табл. 18.3 приведены данные о содержании некоторых промежуточных компонентов обмена углеводов в головном мозге крыс.
Таблица 18.3. Средние данные о содержании некоторых метаболитов обмена углеводов в головном мозге крыс
| Метаболит | Содержание, мкмоль на 1 i сырой массы ткани | Метаболит | Содержание, мкмоль на 1 \ сырой массы ткани |
| Глюкозо-6-фосфат Фруктозо-6-фосфат Фруктозо-1,6-бисфосфа г Диоксиацетонфосфа г Глицеральдегид-3-фосфат | 0,039-0,049 0,017-0,023 0,010-0,017 0,024 0,021-0,046 | З-Фоефоглицерат 2-Фосфоглицерат Фосфоенолиируват Пируват Лактат | 0,085-0,100 0,010-0,016 0,035-0,097 0,120-0,190 1,26-1,70 |
В процентах к общим липидам
| Таблица 18.2 | . Липидный состав нервной ткани | |||
| Серое вещество | Белое вещество | Миелин | ||
| Общее содержание ли-пидов, % от сухой массы | 32,7 | 54,9 |
Адениновые нуклеотиды и креатинфосфат
Из свободных нуклеотидов в мозговой ткани на долю адениновых нуклеотидов приходится около 84%. Большую часть оставшихся нуклеотидов составляют производные гуанина. В целом количество высокоэргических соединений в нервной ткани невелико. Содержание нуклеотидов и креатинфосфата в головном мозге крыс в среднем составляет (в мкмоль на 1 г сырой массы): АТФ — 2,30 — 2,90; АДФ— 0,30-0,50; АМФ- 0,03-0,05; ГТФ - 0,20-0,30; ГДФ - 0,15-0,20; УТФ - 0,17-0,25; креатинфосфат — 3,50 — 4,75. Распределение основных макроэргических соединений примерно одинаково во всех отделах мозга.
Содержание циклических нуклеотидов (цАМФ и цГМФ) в головном мозге значительно выше, чем во многих других тканях. Уровень цАМФ в мозге составляет в среднем 1 — 2, а цГМФ — до 0,2 нмоль на 1 г ткани. Для мозга характерна также высокая активность ферментов метаболизма циклических нуклеотидов. Большинство исследователей считают, что циклические нуклеотиды участвуют в синаптической передаче нервного импульса.
Минеральные вещества
Na, К, Си, Fe, Ca, Mg и Мп распределены в головном мозге относительно равномерно между серым и белым веществом. Содержание фосфора в белом веществе выше, чем в сером.
В табл. 18.4 представлены средние данные о содержании основных минеральных компонентов в головном мозге и плазме крови человека.
Из таблицы 18.4 видно, что
Таблица 18.4. Содержание основных минеральных концентрация ионов калия, компонентов 8 ткани головного мозга и в плазме натрия, а также хлора в мозге крови человека резко отличается от концентра-
ции их в жидкостях тела.
Количественное соотношение неорганических анионов и катионов в мозговой ткани свидетельствует о дефиците анионов. Расчет показывает, что для покрытия дефицита анионов потребовалось бы в 2 раза больше белков, чем их имеется в мозговой ткани. Принято считать,
что остающийся дефицит анионов покрывается за счет липидов. Вполне возможно, что участие липидов в ионном балансе — одна из их функций в деятельности головного мозга.
ОСОБЕННОСТИ МЕТАБОЛИЗМА НЕРВНОЙ ТКАНИ Дыхание
На долю головного мозга приходится 2 — 3 % от массы тела. В то же время потребление кислорода головным мозгом в состоянии физического покоя достигает 20 — 25% от общего потребления его всем организмом, а у детей в возрасте до 4 лет мозг потребляет даже 50% кислорода, утилизируемого всем организмом.
О размерах потребления головным мозгом из протекающей крови различных веществ, в том числе и кислорода, можно судить по артериовенозной разнице. Установлено, что во время прохождения через мозг кровь теряет около 8 об.% кислорода. В 1 мин в расчете на 100 г мозговой ткани протекает 53 — 54 мл крови.
| Компонент | Мозговая ткань, ммоль/кг | Плазма крови, ммоль/л |
| Na+ | ||
| К + | ||
| Са2 + | 2,5 | |
| С1- | ||
| HCOf |
Таблица 18.4 Содержание основных минеральных компонентов 8 ткани головного мозга и в плазме крови человека
Следовательно, 100 г мозга потребляют в 1 мин 3,7 мл кислорода, а весь головной мозг (1500 г) - 55,5 мл кислорода1.
Газообмен мозга значительно выше, чем газообмен других тканей, в частности он превышает газообмен мышечной ткани почти в 20 раз. Интенсивность дыхания для различных областей головного мозга неодинакова. Например, интенсивность дыхания белого вещества в 2 раза ниже, чем серого (правда, в белом веществе меньше клеток). Особенно интенсивно расходуют кислород клетки коры мозга и мозжечка.
Поглощение кислорода головным мозгом значительно уменьшается при наркозе. Напротив, интенсивность дыхания мозга возрастает при увеличении функциональной активности.
Обмен глюкозы и гликогена
Основным субстратом дыхания мозговой ткани является глюкоза, В 1 мин 100 г ткани мозга человека потребляют в среднем 5 мг глюкозы. Подсчитано, что более 90 % утилизируемой глюкозы в ткани мозга окисляются до СО2 и Н2О при участии цикла трикарбоновых кислот. В физиологических условиях роль пенто-.юфосфатного нуги окисления глюкозы в мозговой ткани невелика. Однако этот путь окисления ыюкозы присущ всем клеткам головного мозга. Образующаяся в процессе иеитозофосфатного цикла восстановленная форма НАДФ (НАДФН2) используется на синтез жирных кислот и стероидов.
Интересно отметить, что в расчете на всю массу головного мозга содержание глюкозы в нем составляет около 750 мг. За 1 мин тканью мозга окисляются 75 мг глюкозы. Следовательно, количество глюкозы, имеющееся в ткани головного мозга, могло бы быть достаточным лишь на 10 мин жизни человека. Данный расчет, а также величина артериовенозной разницы но глюкозе доказывают, что основным субстратом дыхания головного мозга является глюкоза крови. По-видимому, глюкоза ле.1 ко диффундирует из крови в ткань головного мозга (содержание глюкозы в мозговой ткани - 0,05 %, а в артериальной крови — 4,44 ммоль/л, или 80 мг/100 мл).
Между глюкозой и гликогеном мозговой ткани имеется тесная связь, выражающаяся в том, что при недостаточном поступлении глюкозы из крови гликоген головного мозга является источником глюкозы, а глюкоза при ее избытке — исходным материалом для синтеза гликогена. Распад гликогена в мозговой ткани происходит путем фосфоролиза с участием системы цАМФ. Однако в целом же использование (ликогена п мозге по сравнению с глюкозой не играет существенной роли в энергетическом отношении, так как содержание гликогена в головном мозге невелико.
Наряду с аэробным метаболизмом уиюводов мозговая ткань способна к довольно интенсивному анаэробному гликолизу. Значение этого явления пока недостаточно ясно, ибо гликолиз как источник энергии ни в коей мере не может сравниваться по эффективности с тканевым дыханием в головном мозге.
Обмен лабильных фосфатов (макроэргов)
Интенсивность обновления богатых энергией фосфорных соединений в головном мозге очень велика. Именно этим можно объяснить, что содержание АТФ и креатин-фосфата в мозговой ткани характеризуется значительным постоянством. При прекращении доступа кислорода мозг может «просуществовать» немногим более минуты за счет резерва лабильных фосфатов. Прекращение доступа кислорода даже на 10—15 с нарушает энергетику нервных клеток, что в целостном организме выражается наступ-
лением обморочного состояния. По-видимому, при кислородном голодании мозг может очень недолго получать энергию за счет процессов гликолиза.
Установлено, что при инсулиновой коме содержание глюкозы в крови может снижаться до 1 ммоль/л, потребление кислорода мозгом в этих условиях не более 1,9 мл/100 г в 1 мин. В норме концентрация глюкозы в крови 3,3 — 5,0 ммоль/л, а мозг потребляет 3,4 — 3,7 мл кислорода на 100 г массы в 1 мин. При инсулиновой коме нарушаются процессы окислительного фосфорилирования в мозговой ткани, снижается концентрация АТФ и происходит изменение функций мозга.
Возбуждение и наркоз быстро сказываются на обмене лабильных фосфатов. В состоянии наркоза наблюдается угнетение дыхания; содержание АТФ и креатин-фосфата повышено, а уровень неорганического фосфата снижен. Следовательно, сокращается потребление мозгом соединений, богатых энергией.
Напротив, при раздражении интенсивность дыхания усиливается в 2 — 4 раза; уровень АТФ и креатинфосфата снижается, а количество неорганического фосфата увеличивается. Эти изменения наступают независимо от того, каким образом произошло стимулирование нервных процессов, а именно: путем электрического разраже-ния или химическим путем.
Обмен белков и аминокислот
Общее содержание аминокислот в ткани мозга человека в 8 раз превышает концентрацию их в крови. Аминокислотный состав мозга отличается определенной специфичностью. Так, концентрация свободной глутаминовой кислоты в мозге выше, чем в любом другом органе млекопитающих (10 мкмоль/г). На долю глутаминовой кислоты вместе с ее амидом глутамином и трипептидом глутатионом приходится более 50 % а-аминоазота головного мозга. В мозге содержится ряд свободных аминокислот, которые лишь в незначительных количествах обнаруживаются в других тканях млекопитающих. Это у-аминомасляная кислота, N-ацетиласпарагиновая кислота и цистатионин (см. главу 11).
Известно, что обмен аминокислот в мозговой ткани протекает в разных направлениях. Прежде всего пул свободных аминокислот используется как источник «сырья» для синтеза белков и биологически активных аминов. Одна из функций дикарбоно-вых аминокислот в головном мозге — связывание аммиака, освобождающегося при возбуждении нервных клеток.
Установлено, что белки в головном мозге находятся в состоянии активного обновления, о чем свидетельствует быстрое включение радиоактивных аминокислот в молекулы белков. Однако в разных отделах головного мозга скорость синтеза и распада белковых молекул неодинакова. Белки cepoi о вещества больших полушарий и белки мозжечка отличаются особенно большой скоростью обновления. Участки головного мозг а, богатые проводниковыми структурами — аксонами (белое вегцество головного мозга), имеют меньшую скорость синтеза и распада белковых молекул.
При различных функциональных состояниях ЦНС наступают изменения в интенсивности обновления белков. Так, при действии на организм животных возбуждающих агентов (фармакологические средства и электрический ток) в головном мозге усиливается интенсивность обмена белков. Напротив, под влиянием наркоза скорость распада и синтеза белков снижается.
Возбуждение нервной системы сопровождается повышением содержания аммиака в нервной ткани. Это явление наблюдается как при раздражении периферических нервов, так и при раздражении мозга. Считают, что образование аммиака при возбуждении в первую очередь происходит за счет дезаминирования АМФ.
Аммиак — очень ядовитое вещество, особенно для нервной системы. Особую роль в устранении аммиака играет глутаминовая кислота. Она способна связывать
аммиак с образованием глутамина — безвредного для нервной ткани вещества. Данная реакция амидирования протекает при участии фермента глутаминсинтетазы и требует затраты энергии АТФ (см. главу 11). Непосредственный источник глутаминовой кислоты в мозговой ткани — путь восстановительного аминирования ос-кетоглутаровой кислоты:
Образование глутаминовой кислоты из а-кетоглутаровой и аммиака является важным механизмом нейтрализации аммиака в ткани мозга, где путь устранения аммиака за счет синтеза мочевины не играет существенной роли.
Кроме того, глутаминовая кислота образуется и в процессе переаминирования. Активность АсАТ в мозговой ткани значительно выше, чем в печени и особенно в почках.
Наконец, глутаминовая кислота в нервной ткани может декарбоксилироваться с образованием ГАМК:
ГАМК в наибольшем количестве содержится в сером веществе головного мозга. В спинном мозге и периферических нервах ее значительно меньше.
Обмен липидов
Липиды составляют около половины сухой массы головного мозга. Как уже отмечалось, в нервных клетках серого вещества особенно много фосфоглицеридов, а в мнелиновых оболочках нервных стволов — сфингомиелина. Из фосфоглицеридов серого вещества мозга наиболее интенсивно обновляются фосфатидилхолины и особенно фосфатидилинозитол. Обмен липидов миелиновых оболочек протекает с небольшой скоростью. Холестерин, цереброзиды и сфингомиелины обновляются очень медленно.
Ткань головного мозга взрослого человека содержит много холестерина (около 25 г). У новорожденных в головном мозге всего 2 г холестерина; количество его резко возрастает в первый год жизни (примерно в 3 раза). При этом биосинтез холестерина происходит в самой мозговой ткани. У взрослых же людей синтез холестерина в головном мозге резко снижается, вплоть до полного прекращения.
ХИМИЧЕСКИЕ ОСНОВЫ ВОЗНИКНОВЕНИЯ И ПРОВЕДЕНИЯ НЕРВНЫХ ИМПУЛЬСОВ
Каковы же химические основы возникновения и поддержания биоэлектрических потенциалов (потенциалов покоя и действия)? Большинство исследователей придерживаются мнения, что явления электрической поляризации клетки обусловлены неравномерным распределением ионов калия и натрия по обе стороны клеточной мембраны. Мембрана обладает избирательной проницаемостью: большей для ионов калия и значительно меньшей для ионов натрия. Кроме того, в нервных клетках существует механизм, который поддерживает внутриклеточное содержание натрия на низком уровне вопреки градиенту концентрации. Этот механизм получил название «натриевого насос а».
При определенных условиях резко повышается проницаемость мембраны для ионов натрия.
В состоянии покоя внутренняя сторона клеточной мембраны заряжена электроотрицательно по отношению к наружной поверхности. Объясняется это тем. что количество ионов натрия, выкачиваемых из клетки с помощью натриевого насоса, не вполне точно уравновешивается поступлением в клетку ионов калия. 13 связи с этим часть катионов натрия удерживается внутренним слоем противоионов (анионов) на наружной поверхности клеточной мембраны.
При возбуждении, вызванном тем или иным агентом, селективно изменяется проницаемость мембраны нервной клетки (аксона): увеличивается избирательно для ионов натрия (примерно в 500 раз) и остается без изменения для ионов калия. В результате ионы натрия устремляются внутрь клетки. Компенсирующий поток ионов калия, направляющийся наружу из клетки, несколько запаздывает1. Это приводит к возникновению отрицательного заряда на наружной поверхности клеточной мембраны. Внутренняя поверхность мембраны приобретает положительный заряд; происходит перезарядка клеточной мембраны (в частности, мембраны аксона, т. е. нервного волокна) и возникает потенциал действия, или спайк. Продолжительность спайка не превышает 1 мс. Он имеет восходящую фазу, пик и нисходящую фазу. Нисходящая фаза (падение потенциала) связана с нарастающим преобладанием выхода ионов калия над поступлением ионов натрия — мембранный потенциал возвращается к норме. После проведения импульса в клетке восстанавливается состояние покоя. В этот период ионы натрия, вошедшие в нейрон при возбуждении, заменяются на ионы калия. Этот переход происходит против фадиента концентрации, так как ионов натрия во внешней среде, окружающей нейроны, намного больше, чем в клетке после момента ее возбуждения. Переход ионов натрия против градиента концентрации, как уже отмечалось, осуществляется с помощью натриевого насоса, для работы которого необходима энергия АТФ. В конце концов все это приводит к восстановлению исходной концентрации катионов калия и натрия внутри клетки (аксона), и нерв готов для получения следующего импульса возбуждения. Другим не менее важным процессом для нервной ткани является передача нервного импульса от одной нервной клетки к другой или воздействие на клетки эффектор-ного органа.
Роль медиаторов в передаче нервных импульсов
Связь миллиардов нейронов мозга осуществляется посредством.медиаторов. Химическое вещество можно отнести к числу медиаторов лишь в том случае, если оно удовлетворяет ряду критериев. В нервных волокнах должны содержаться ферменты, необходимые для синтеза этого вещества. При раздражении нервов это вещество должно выделяться, реагировать со специфическим рецептором на постси-наптической клетке и вызывать биологическую реакцию. Должны существовать механизмы, быстро прекращающие действие этого химического вещества.
Всем этим критериям удовлетворяют два вещества — ацетилхолин и норадреналин. Содержащие их нервы называют соответственно холинергическими и адренерги-ческими. В соответствии с этим все эфферентные системы делят на холиноре-цепторы и адренорецепторы.
Ряд других химических веществ удовлетворяет многим, но не всем перечисленным критериям. К таким медиаторам относятся дофамин, адреналин, серотонин, октопамин, гистамин, ГАМК и др.
Обширная группа холинорецепторов весьма неоднородна как в структурном, так и в функциональном отношении. Объединяют их медиатор — ацетилхолин — и общая схема строения синапса.
Ацетилхолин представляет собой сложный эфир уксусной кислоты и холиыа. Он синтезируется в нервной клетке из холина и активной формы ацетата — аце-
тилкоэнзима А при помощи специального фермента холинацетилтрансферазы (холйн-ацетилазы):
Синапс можно представить себе как узкое пространство (щель), ограниченное с одной стороны пресинаптической, с другой — постсинаптической мембраной (рис. 18.4). Пресинапти-ческая мембрана состоит из внутреннего слоя, принадлежащего цитоплазме нервного окончания, и наружного слоя, образованного нейроглией. Мембрана в некоторых местах утолщена и уплотнена, в других -- истончена и имеет отверстия, с помощью которых цитоплазма аксона может сообщаться с синаптическим пространством. Постсинаптическая мембрана менее плотная, не имеет отверстий. Подобным образом построены и нервно-мышечные синапсы, но они имеют более сложное строение мембранного комплекса.
В общих чертах картину участия ацетилхолина в осуществлении передачи нервного возбуждения можно представить следующим образом. В синаптических нервных окончаниях имеются пузырьки (везикулы) диаметром 30 — 80 нм, которые содержат нейромедиаторы. Эти пузырьки покрыты оболочкой, которая образована белком клатрином (молекулярная масса 180000 Да). В холи-нергических синапсах каждый пузырек диаметром 80 нм содержит ~ 40 000 молекул ацетилхолина. При возбуждении высвобождение медиатора происходит «квантами», т. е. путем полного опорожнения каждого отдельного пузырька. В нормальных условиях под влиянием сильного импульса выделяется примерно 100 — 200 квантов медиатора — количество, достаточное для инициирования потенциала действия в постсинаптическом нейроне. Происходит это, по-видимому, так: деполяризация мембраны синаптических окончаний вызывает быстрый ток ионов кальция в клетку. Временное увеличение внутриклеточной концентрации ионов кальция стимулирует слияние мембраны синаптических пузырьков с плазматической мембраной и таким образом запускает процесс высвобождения их содержимого. Для выброса содержимого одного пузырька требуются примерно четыре иона кальция. Выделенный в синаптическую щель ацетилхолин вступает во взаимодействие с белком-хеморецептором, входящим в состав постсинаптической мембраны. В результате изменяется проницаемость мембраны — резко увеличивается ее пропускная способность для ионов натрия. Взаимодействие между
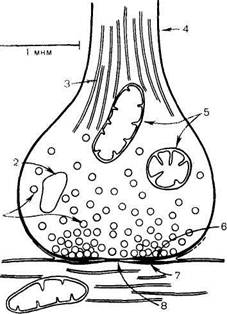
Рис. 18.4. Схематическое изображение синапса (по Мецлеру).
1 — синаптические пузырьки; 2 — лизосома; 3 — микрофибриллы (нейрофибриллы); 4 —аксон; 5 — митохондрии; 6 — пресинаптическое утолщение мембраны; 7 — постеинаптическое утолщение мембраны; 8 — синаптическая щель (около 20 нм).
рецептором и медиатором запускает ряд реакций, заставляющих постсинапти-ческую нервную клетку или эффекторную клетку выполнять свою специфическую функцию. После выделения медиатора должна наступить фаза его быстрой инактивации или удаления, чтобы подготовить синапс к восприятию нового импульса. В холинергических синапсах это происходит двумя путями. Прежде всего ацетил-холин подвергается ферментативному гидролизу. Второй путь — это энергозависимый активный транспорт ацетилхолина в нейрон, где он накапливается для последующего повторного использования.
Гидролитический распад ацетилхолина на уксусную кислоту и холин катализируется ферментом, который получил название ацетилхолинэстеразы:
В большинстве отделов головного мозга гидролиз ацетилхолина осуществляется ацетил-холинэстеразой (истинной холинэстеразой, которая гидролизует ацетилхолин быстрее, чем иные эфиры холина). В нервной ткани существую! и другие эстеразы, которые способны гидроли-зовать ацетилхолин, но значительно медленнее, чем, например, бутирилхолин. Эти эстеразы называются холинэстеразой (или псевдохолинэстеразой). К числу холинергических систем относятся моторные нейроны, образующие нервно-мышечные соединения, все преганглионарные нейроны автономной нервной системы и постгаиглионарные нейроны парасимпатической нервной системы. Большое количество холинергических симпатических областей обнаружено также в головном мозге. В зависимости от чувствительности к той или иной группе химических соединений холинергические нейроны делятся на «мускариновые» (активируемые мускари-ном) и «никотиновые» (активируемые никотином). Мускариновые рецепторы ацетилхолина, имеющиеся во многих нейронах автономной системы, специфически блокируются атропином. Никотиновые синапсы присутствуют в ганглиях и скелетных мышцах. Их ингибиторами являются кураре и активный компонент этого яда — D-тубокурарин,
Необходимо подчеркнуть, что в адренорецепторах существует два вида рецепторов для норадреналина: а- и Р-адренергические рецепторы. Эти рецепторы можно отличить друг от друга по специфическим реакциям, которые они вызывают, а также по тем специфическим агентам, которые способны блокировать данные реакции.
Р-Адренергические рецепторы включают эфферентную клетку с помощью аде-нозин-3', 5'-монофосфата, или цАМФ — универсального «второго посредника» между гормонами и различными функциями клеток, на которые воздействуют гормоны (см. главу 6).
Установлено, что как только Р-адренергический рецептор (расположенный на наружной поверхности мембраны эффекторной клетки) начнет взаимодействовать с норадреналином, на внутренней поверхности клеточной мембраны активируется фермент аденилатциклаза. Затем в клетке аденилатциклаза превращает АТФ в цАМФ; последний в свою очередь способен оказывать влияние на метаболизм клетки. Этот сложный ряд последовательных реакций может быть заблокирован пропранололом — веществом, препятствующим связыванию норадреналина с р-адре-нергическим рецептором.
Известно, что в метаболизме катехоламиновых медиаторов особая роль принадлежит ферменту моноаминоксидазе (МАО). Этот фермент удаляет аминогруппу (— NH2) у норадреналина, серотонина, дофамина и адреналина, тем самым инактивируя упомянутые медиаторы. Однако в последние годы было показано, что, помимо ферментативного превращения, существует и другой механизм быстрой инактивации, точнее удаления, медиаторов. Оказалось, что норадреналин быстро исчезает из си-
наптической щели в результате вторичного поглощения симпатическими нервами; вновь оказавшись в нервном волокне, медиатор, естественно, не может воздействовать на постсинаптические клетки. Конкретный механизм этого явления пока не вполне ясен.
Адренергическая и холинергическая системы головного мозга тесно взаимодействуют с другими системами мозга, в частности использующими серотонин в качестве медиатора. В основном серотонинсодержащие нейроны сосредоточены в ядрах мозгового ствола. Нейромедиаторная роль серотонина осуществляется в результате взаимодействия серотонина со специфическими серотонинергическими рецепторами. Исследования, проведенные с ингибитором синтеза серотонина — и-хлорфенилаланином, а также с другими ингибиторами, дают основания считать, что серотонин влияет на процессы сна. Выявлено также, что торможение кортикостероидами секреторной активности гипофиза оказывается менее эффективным у тех животных, мозг которых беднее серотонином.
Важным нейромедиатором, выполняющим тормозные функции, является ГАМ К, количество которой в головном мозге во много раз выше, чем других нейромедиа-торов. Так, в гипоталамусе суммарное содержание ацетилхолина, норадреналина, дофамина и серотонина не превышает 10 мкг/г, в то время как ГАМК в этом отделе головного мозга более 600 мкг/г.
В настоящее время в терапевтической практике применяется большое количество лекарственных средств, которые действуют через систему медиаторов. Многие лекарственные вещества, успешно применяемые при лечении гипертонии, влияют на накопление и выделение адренергических медиаторов. Например, резерпин — понижающее артериальное давление средство — специфически тормозит процесс переноса катехоламинов в специальные гранулы нейронов и тем самым делает эти амины доступными действию эндогенной МАО.
Гипотензивные лекарственные препараты, такие как а-метилдофа, под действием содержащихся в нервной клетке (аксоне) ферментов превращаются в вещества, напоминающие по своему строению норадреналин. Эти «ложные» медиаторы накапливаются и выделяются вместе с естественными медиаторами, разбавляя их и тем самым снижая их эффект.
Многие антидепрессанты (вещества, снимающие депрессию) увеличивают содержание катехоламинов в синаптической щели, т. е. количество медиаторов для стимулирования рецептора возрастает. К таким веществам, в частности, относятся имипрамин (блокирует поглощение норадреналина нервными волокнами), амфетамин (одновременно способствует выделению норадреналина и блокирует его поглощение), ингибиторы МАО (подавляют метаболизм катехоламинов) и др. В связи с этим возникла катехоламиновая гипотеза депрессивных состояний, согласно которой психическая депрессия связана с недостатком катехоламинов в мозге.
В начале 50-х годов фармакологи выяснили, что известный галлюциноген — диэтиламин лизергиновой кислоты (ЛСД) не только сходен по химическому строению с серотонином, но и нейтрализует некоторые его фармакологические эффекты (блокируя рецепторы серотонина). Поэтому было высказано предположение, что нарушение обмена серотонина может быть причиной возникновения особых психических заболеваний.
Считают, что такие антипсихотические средства, как аминазин (хлорпромазин) и галопе-ридол, усиливая синтез катехоламинов, способны блокировать дофаминовые рецепторы в мозге.
Механизмы памяти
Память не сосредоточена в одном строго локализованном участке мозга, подобно центрам зрения, слуха, речи и т. д. В то же время память — не свойство всего мозга в целом. Субстратом памяти человека являются нейроны.
Память человека нельзя рассматривать в отрыве от его деятельности, так как не познание познает, не мышление мыслит, не память запоминает и воспроизводит, а познает, мыслит, запоминает и воспроизводит человек, определенная личность.
В последние годы отчетливо показано, что обучение животного новым навыкам отражается на химизме мозговых клеток (нейронов): меняются количество уридина в цитоплазматической РНК, степень метилирования ДНК и фосфорилирования
ядерных белков. Применение стимуляторов и веществ — предшественников РНК облегчает обучение, а введение блокаторов синтеза РНК, напротив, затрудняет этот процесс. Существуют данные, что после запоминания информации меняется антигенный состав мозговой ткани. Несомненно, память — цепь процессов, в которых сложные вещества, в частности РНП и в первую очередь информосомы, играют существенную роль. Принято выделять несколько форм биологической памяти: генетическую, иммунологическую и нейрологическую.
Биохимические основы генетической памяти более или менее ясны. Ее носителем является ДНК клетки. Следующей по сложности формой памяти является иммунологическая. Этот вид памяти хотя и включает элементы генетической памяти, но находится на более высокой ступени сложности. Наконец, система нейрологической памяти еще более сложна. Эта форма в свою очередь может быть разделена на кратковременную память (КП) и долговременную память (ДП). В основе КП, по всей вероятности, лежит «циркуляция» информации, полученной в виде импульсов, по замкнутым цепям нейронов. При этом синаптический эффект, изменения ядерно-ядрышкового аппарата, выброс в цитоплазму нейрона биологически активных веществ и сопутствующая этим процессам перестройка обмена веществ клетки — все это может расцениваться как показатели функционирования КП.
Включение блоков ДП обеспечивается примерно через 10 мин после прихода информации в клетку. За это время происходит перестройка биологических свойств нервной клетки. Ряд исследователей считают, что афферентная импульсация, приходящая в нервные клетки во время обучения, вызывает либо чисто количественную активацию синтеза РНК и белка, что может приводить к установлению новых синаптических связей и перестройке существующих, либо наступающая активация синтеза нуклеиновых кислот и белка носит целенаправленный, специфический характер, а синтезированные молекулы являются хранилищем информации.
Пептиды и болевые реакции
В 70-х годах в головном мозге различных позвоночных животных были обнаружены специфические рецепторы морфина. Эти рецепторы сосредоточены на синаптических мембранах, наиболее богата ими лимбическая система, от которой зависит эмоциональный ответ. В дальнейшем из мозговой ткани выделили эндогенные пептиды, имитирующие при инъекциях различные эффекты морфина. Эти пептиды, обладающие способностью специфически связываться с опиатными рецепторами, получили название эндорфинов и энкефалинов (см. главу 6).
Оказалось, что пептиды с морфиноподобной активностью являются производными Р-липотропного гормона гипофиза. Установлено, что Р-эндорфин представляет собой фрагмент р-липотропина с 61-го по 91-й, у-эндорфин — с 61-го по 77-й и ос-эндорфин — с 61-го по 76-й аминокислотный остаток.
Энкефалины — также фрагменты Р-липотропина, но они значительно меньше, чем эндорфины. Энкефалины являются пентапептидами. Наиболее изучены два пентапеп-тида: метионинэнкефалин (Тир — Гли — Гли — Фен — Мет) и лейцинэнкефалин (Тир-Гли-Гли-Фен-Лей). Содержание метионинэнкефалинов в головном мозге в 4 раза превышает содержание лейцинэнкефалинов.
|
|
|
|
|
Дата добавления: 2014-12-27; Просмотров: 2958; Нарушение авторских прав?; Мы поможем в написании вашей работы!