КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Основные этапы филогенетического развития животного мира 1 страница
|
|
|
|
Трубку
Второй сегмент, S — третий сегмент (туловище), 7— спинная ресничная полоска. 8 — мужское половое отверстие, 9 — просвечивающие железы, выделяющие
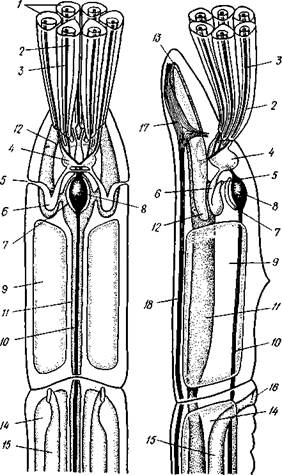
телиальную клетку, содержит одно ядро и длинную петлю тончайшего капиллярного сосуда. Кровь попадает в этот капилляр из приносящего сосуда щупальца, а выносится из него в выносящий щупальцевый сосуд. Таким образом, межщупальцевое пространство играет роль своеобразной «кишечной полости», пиннулы — «кишечных ворсинок», а сами по-гонофоры представляют совершенно исключительный пример свободно-живущих животных, у которых обычное кишечное пищеварение полностью заменено наружным пищеварением.
Кровеносная система замкнутая, состоит в основном из спинного и брюшного сосудов (рис. 563). Спинной сосуд способен сокращаться и гонит кровь сзади наперед; у основания щупалец он расширяется, образуя мускулистое сердце, к которому иногда прилегает эпителиальный пузырек перикардия, напоминающий таковой гемихордовых. Приносящие сосуды щупалец берут начало от спинного сосуда, брюшной сосуд образуется слиянием выносящих щупальцевых сосудов. Кровь окрашена в красный цвет растворенным в ней гемоглобином.
Органы выделения представлены парой сильно развитых почек, открывающихся внутренними концами в передний непарный целом; они
представляют собой целомодукты 1-го сегмента (рис. 563). В отличие от гемихордовых 2-й сегмент погонофор без целомодуктов. В 3-м, туловищном, сегменте имеется своя пара целомодуктов, но здесь они играют роль половых протоков.
|
Нервная система очень примитивна и целиком залегает в толще кожного эпителия, напоминая нервную систему Hemichordata. В эпителии головной лопасти находится мозг — брюшное скопление ганглиозных клеток. От мозга назад тянется брюшной нервный тяж и берут начало нервы щупалец. На брюхе туловищного отдела имеется полоска мерцательного эпителия; возможно, это орган химического чувства.
Половая система. Пого-нофоры раздельнополы, половой диморфизм отсутствует. Половая система лежит в 3-м сегменте. У самцов в задней половине туловища помещается пара длинных колбасовидных семенных мешков, открывающихся посредством длинных целомодуктов — семяпрово-водов на передней границе туловища. В семяпроводах формируются мешковидные сперматофоры, содержащие сперматозоиды и снабженные длинной нитью. У самок пара вытянутых яичников лежит в передней части туловища, а сравнительно короткие яйцеводы открываются в средней части туловища.
Развитие. Самка откладывает яйца в передний участок собственной трубки, где протекает все развитие. Яйца более или менее удлиненные, богатые желтком.
| Рис.563. Схема строения Pogonophora (по А. Ива-нону). А — передний участок тела самца со спин-нон стороны; Б — то же, с левой стороны: 1 —• щупальца, 2 — цсломический канал щупальца,,? — приносящий и уносящий сосуды щупальца, 4 — целом первого сегмента, 5 — наружное отверстие целомодукта, 6 — целомодукт первого сегмента, 7 — перикардий, 8 — сердце, 9 -- целом второго сегмента, 10 — спинной сосуд, 11 — брюшной сосуд, 12 — боковой головной сосуд, 13 — срединный головной сосуд, 14 — целом третьего сегмента, 15 —семяпровод, 16 — половое отверстие, 17 — мозг, 18 — брюшной нервный ствол |
Дробление яйца напоминает спиральное, детерминированное. Однако в отличие от кольчатых червей и моллюсков у погонофор целоми-ческая мезодерма образуется не из телобластов 4d, a за счет потомков переднего макромера В. В результате дробления образуется плотный зародыш. Мелкие клетки эктодермы постепенно обрастают крупные энтодер-мальные бластомеры, остав-
|
ляя на будущей брюшной стороне небольшое отверстие бластопора. В зачатке энтодермы появляется полость, и он превращается в замкнутый со всех сторон первичный кишечник. От последнего спереди отделяется пара мезодермальных мешков — зачатки целома, которые быстро растут в заднем направлении. После этого тело зародыша расчленяется на отделы, причем сначала обособляется четвертый, самый задний первичный сегмент, или отдел тела, и в него попадают задние участки целома (рис. 564). Затем формируется третий сегмент с его целомически-ми мешками, наконец, появляется граница между целомами первого и второго сегментов.
Сначала развивается одно левое щупальце, в него заходит продолжение левого переднего целомического мешочка, потом одно за другим вырастают остальные щупальца. Все они сообщаются с левым передним целомом. Околосердечная сумка (перикардий) образуется из правого переднего целомического мешочка.
Итак, целом первичных сег- рис. 564. Схема развития целома и расчлене-
ментов у Pogonophora образу- ние тела у погонофор (по А. В. Иванову). А —
ется так же как И у ИГЛОКОЖИХ появление энтероцельных выпячиваний кишеч-
и китпрчнп'пытпятпих (с ^W ника; 5 —Р°ст энтероцельных целомических
И кишечнодышащих (С. ООЭ), мешков; S — отделение задней пары целомиче-
т. е. знтероцельным путем, ских мешков и заднего сегмента (телосомы);
Внутренняя масса энтодер- Г —отделение от передних целомов метасомы; мальных клеток, оставшихся Д — развитие 1-го щупальца и появление ще-после отделения целома, пред- тинок на телосоме; Е- закладка 2-го щупаль-
„ г ца, отделение от передних целомов мезоцелеи;
ставляет зачаток средней киш- ж-образование септ в задних целомах (те
ки, однако далее ОН не разви- лосоме) и формирование непарного переднего
вается и в конце эмбриональ- целома (протоцеля)
ного развития рассасывается.
В дальнейшем на переднем и
заднем концах зародыша развиваются ресничные пояски, но они временные и у взрослого животного не сохраняются. Молодое животное выходит из материнской трубки и сразу же переходит к жизни на дне, выделяя собственную трубку. Дальнейшее развитие сводится к усиленному росту, образованию прикрепительных органов, половых органов и т. д. Свободноплавающая личинка в развитии изученных до сих пор в эмбриологическом отношении погонофор отсутствует.
Филогения. Образ жизни погонофор сходен с таковым сидячих поли-хет. Не удивительно поэтому, что и внешний вид, и даже некоторые детали наружного строения в этих двух группах обнаруживают много общего. Конвергентное поверхностное сходство послужило некоторым зоологам поводом неправильно рассматривать погонофор как аберрантных многощетинковых кольчецов из подкласса Sedentaria. В организации погонофор мы видим много характерных особенностей, присущих также аннелидам, эхиуридам, моллюскам и отчасти щупальцевым и ше-• тинкочелюстным. Это, с одной стороны, способность образовывать хитин
и щетинки «аннелидного типа». С другой стороны, погонофоры сходны с частью щупальцевых (плеченогими), с щетинкочелюстными и с вто-ричноротыми энтероцельным способом образования вторичной полости тела, а также асимметрическим развитием передней пары целомиче-ских мешков, образованием перикардия из правого переднего целома.
Подобно аннелидам и хордовым, погонофоры — животные сегментированные и полимерные (состоящие из многих сегментов). Однако сегментация в этих трех группах развилась независимо и различными путями. У погонофор наиболее отчетливо сегментирован задний отдел тела, и это объясняется особенностями их образа жизни. Дело в том, что погонофора в течение жизни почти непрерывно закапывается в грунт задним концом, который приспособлен для активного рытья и выдвигается для этого через заднее отверстие трубки. Метамерные щетинки и мускулистые поперечные перегородки, разделяющие целом заднего отдела тела, развились как необходимые приспособления к активному рытью. По мере зарывания животного в ил трубка сзади постепенно удлиняется.
Вероятно, эволюция погонофор пошла по пути глубокого приспособления к сидячему образу жизни в защитной трубке, причем одним из своеобразных процессов явилась редукция кишечника и физиологическая замена его щупальцевым аппаратом.
Эволюция погонофор сопровождалась множественной закладкой прикрепительных хитиноидных пластиночек и вторичным упорядочиванием их расположения. Щупальцевый аппарат в одних случаях испытал оли-гомеризацию (единственное щупальце Siboglinum), в других, напротив, количество щупалец значительно возросло (Lamellisabella, Spirobrachia и Lamellibrachia).
ТИП ЩЕТИНКОЧЕЛЮСТНЫЕ (CHAETOGNATHA)
Щетинкочелюстные — небольшая, своеобразная, довольно обособленная от других типов группа морских планктонных животных. Наиболее вероятно их родство с Deuterostomia, на что особенно указывает их эмбриональное развитие.
1. Тело щетинкочелюстных состоит из трех отделов — головы, туловища и хвоста, причем каждый отдел имеет собственный участок целома.
2. На голове расположены две боковые группы мощных крючкооб-разных хватательных щетинок.
3. Тело окаймлено плавниками — парными боковыми и хвостовым.
4. Нервная система состоит из надглоточного и подглоточного ганглиев, соединенных окологлоточными коннективами, и подкожного нервного сплетения.
5. Кровеносная и выделительная системы отсутствуют.
6. Во время эмбрионального развития целом образуется энтероцельным путем.
К типу Chaetognatha принадлежит только один класс с тем же названием.
КЛАСС ЩЕТИНКОЧЕЛЮСТНЫЕ (CHAETOGNATHA)
Известно всего 50 видов щетинкочелюстных, большая часть которых в течение всей жизни обитает в планктоне. Однако представители рода Spadella ведут донный образ жизни. Щетинкочелюстные— быстрые, плавающие как бы толчками (вследствие характерных изгибаний тела в спиннобрюшном направлении) хищники, поедающие планктонных
животных и нападающие порой на добычу, превосходящую их размерами.
Тело щетинкочелюстных стекловиднопрозрачно; по наличию плавников и по характеру движения они несколько напоминают маленьких рыбок. Размеры тела колеблются от 1 до нескольких сантиметров, Sagiita gazellae имеет 10 см в длину.
Строение. Удлиненное тело несколько сужается к концам (рис. 564). На заднем конце и по бокам тела кожа образует выдающиеся складки: хвостовой и боковые плавники. Тело явственно делится двумя поперечными перегородками на три отдела: головной, туловищный и хвостовой. Головной конец прикрыт сверху и с боков двойной кожной складкой — капюшоном. Тело покрыто многослойным кожным эпителием (редкий случай среди беспозвоночных), под которым имеется слой продольных мышц, имеющих поперечнополосатое строение. Мускулатура подстилается изнутри перитонеальным эпителием, ограничивающим обширный целом.
Кишечник в виде прямой трубки; начинается ртом на переднем конце тела и кончается порошицей на границе между туловищем и хвостом. Рот окружен с боков двумя рядами хитиновых подвижных крючков, отчасти прикрытых капюшоном. Кишечник подвешен на спинно-брюш-ном мезентерии, делящем полость тела на правую и левую половины. Chaetognatha — хищники.
Нервная система состоит из надглоточного и подглоточного ганглиев, образующих мозг, и из мощного брюшного узла, лежащего в туловище под кишечником. Мозг и брюшной узел соединены длинными коннек-тивами; брюшной ганглий дает 12 пар нервов к стенке тела, где они соединяются с хорошо развитым подкожным нервным сплетением.
Над мозгом лежит пара маленьких глаз, причем каждый из них состоит из трех слившихся глазков, о чем можно судить по наличию трех хрусталиков. Глаза инвертированы. Позади глаз, на затылке, имеется площадка мерцательного эпителия, которой придают значение органа обоняния. Кроме того, по телу рассеяно значительное количество осязательных эпителиальных бугорков, усаженных на вершине чувствительными волосками.
Половая система. Chaetognatha — гермафродиты. Женские половые органы помещаются в задней части туловища, мужские — в хвостовом отделе. По бокам задней кишки лежат два яичника, от них отходят яйцеводы, открывающиеся наружу на боковых стенках тела. Интересно, что внутренние, обращенные к яичникам концы яйцеводов замкнуты слепо, так что сообщение с яичниками устанавливается, вероятно, лишь в период половой деятельности.
В каждом из боковых отделов хвостовой полости тела имеется по массивному клеточному валику, семеннику, прилегающему к стенке тела. Созревающие живчики попадают в целом и выводятся наружу через два семяпровода, которые начинаются широкой воронкой и открываются наружу по бокам хвостового отдела. Каждый семяпровод образует на своем пути расширение — семенной пузырь, в котором скапливаются живчики. Размножение исключительно половое.
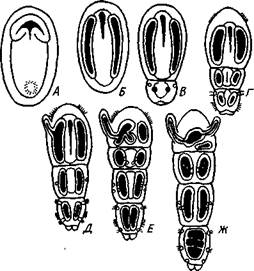
Развитие прямое. Полное равномерное дробление приводит к образованию бластулы, а затем инвагинационной гаструлы. Зачатки половых желез закладываются на стадии 32 бластомеров (рис. 565). Еще в неоплодотворенном яйце имеется небольшое скопление особых зерен, называемое половым детерминантом. На стадии 32 бластомеров детерминант оказывается в одной из клеток. Дальнейшее деление этой клетки происходит так, что одна из дочерних клеток получает целиком весь де-
терминант и становится половым зачатком, тогда как другая лишена детерминанта и дает впоследствии всю энтодерму. Половой бластомер сначала делится надвое, а в гаструле половой зачаток имеет вид двух пар клеток, причем из одной пары возникают в дальнейшем яичники, из другой — семенники. Далее следует отметить энтероцельное образование мезодермы, отшнуровываюшейся от кишечника в виде двух боковых выпячиваний, полость которых представляет целом. Получаются два цело-мических мешка, из которых каждый перешнуровывается поперек на маленький головной и большой туловищный целомы. Последние позднее отделяют хвостовой целом. Бластопор гаструлы замыкается, и окончательно рот возникает независимо от него на противоположном конце зародыша.
Филогения щетинкочелюстных. Тип Chaetognatha относится к вторич-норотым по способу закладки мезодермы и характеру возникновения рта. Однако тело щетинкочелюстных, судя по характеру его развития, слагается из двух сегментов, ибо перегородка, отделяющая полость хвоста от полости туловища, возникает значительно позже и носит вторичный характер. Этим тип Chaetognatha существенно отличается от вто-ричноротых, которые в основном трехсегментные животные.
Таким образом, происхождение щетинкочелюстных еще совершенно не ясно и положение их в системе животных неопределенно. П
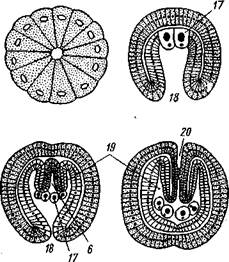
Рис. 565. Схема строения (А — вид с брюшной стороны, по Деляжу) и развития (Б — четыре последовательные стадии, по Р. Гертвигу) Sagitta: I — мозг, 2 — окологлоточный коннектив, 3 — брюшной ганглий, 4 — нервы, 5 — целом, 6 — кишка, 7 — яичник, 8 — яйцевод, 9 — анус, 10 — семенник, // — семяпровод, 12 — хвостовой плавник, 13 — головная железа, 14— хи-тиноидные крючки — щетинки, 15 — мужское половое отверстие, 16 — женское половое отверстие, 17 — первичные половые клетки, 18 — бластопор, 19 — боковые выпячивания первичной кишки — зэчатки целома (энтероцельный способ образования целома), 20 — вторичный рот
В настоящей главе мы дадим краткий обзор усложнения организации беспозвоночных, возникающего в процессе эволюции, не останавливаясь на родственных отношениях отдельных групп. Филогения отдельных групп кратко разобрана в соответствующих местах учебника.
Рассматривая огромное разнообразие форм беспозвоночных, можно по степени сложности строения расположить эти формы на нескольких уровнях. В главных чертах это ярусное расположение отвечает эволюционному пути, пройденному животными. На филогенетическом древе ярусы могут быть отделены друг от друга горизонтальными линиями. Группы животных, расположенные между одними и теми же горизонталями, имеют приблизительно одинаковую сложность организации (см. табл. на с. 578).
Первый этаж родословного древа образован полцарством Protozoa, включающим пять типов. Мы уже видели, что в разных типах Protozoa (Sarcomastigophora и др.) обнаруживается тенденция к переходу в многоклеточное состояние (многоядерность, образование колоний и т. п.). Эта тенденция большей частью остается незавершенной, но у каких-то Mastigophora привела к прочному переходу к многоклеточное™: возникли первые Metazoa.
Низшие Metazoa образуют следующие этажи родословного древа. К ним принадлежат четыре типа Placozoa: Spongia, Coelenterata и Cte-nophora. Из них губки находятся на гораздо более низком уровне развития, о чем прежде всего свидетельствует отсутствие у них настоящих тканей, а также нервной системы. Кроме того, губки существенно отличаются от других Metazoa «извращением» положения зародышевых листков. В связи с этим следует считать, что Spongia отделились от общего ствола Metazoa чрезвычайно рано, вероятно еще на уровне фаго-цителлы — гипотетического организма, который, по мнению И. И. Мечникова, был общим предком всех многоклеточных животных. Таким образом, губки составляют среди Metazoa особый ярус — надраздел Ра-razoa, противопоставляемый всем остальным многоклеточным — надраз-делу Eumetazoa. Coelenterata и Ctenophora образуют следующий ярус — раздел Radiata. Общность особенностей строения позволяет эти два типа именовать Radiata или Diploblastica. Характеризуются они тем, что тело их состоит из производных двух зародышевых листков — эктодермы и энтодермы, которые даже у взрослых животных сохраняют свою обособленность и характерное расположение. По плану строения они сходные гаструлой и обладают радиальной симметрией. Можно предположить, что первичные Coelenterata произошли от фагоцителлы, причем Ctenophora сохранили древний способ движения — плавание посредством ресничного аппарата, a Coelenterata перешли к сидячему образу жизни. В обоих случаях сохранилась первобытная радиальная симметрия тела.
20—763 577
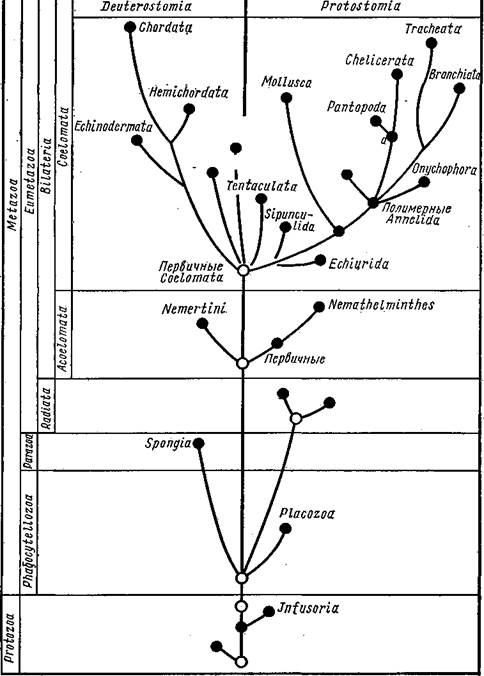
|
| Chaetognatha Poaonophora. |
| Trilobitomorph Tardlgrada |
| 0/шгомернь/е Annelida |
| Plathelmintties |
| ацелооУразные Bilaterlu |
| Ctenophora Coelenteraia |
| Фагоцителла |
| Колониальные Mastigoptiora. Mastigophora Sarcodim |
| Первичные эукариотные простейшие |
Все вышестоящие Eumetazoa в отличие от Radiata характеризуются билатеральной симметрией тела и объединяются в раздел Bilateria. Другое их название Triploblastica, так как тело их формируется в онтогенезе из трех зародышевых листков — экто-, эндо- и мезодермы.
Ни губки, ни кишечнополостные не дали начала новым типам животных. По-видимому, Bilateria произошли независимо от тех и других, непосредственно от фагоцителлообразных предков, перешедших к ползающему образу жизни, вызвавшему появление билатеральной симметрии. Наиболее примитивные Bilateria в мире животных представлены свободноживущими плоскими червями — Plathelminthes, именно классом ресничных червей Turbellaria. Среди последних наиболее примитивные представители — отряд бескишечных Acoela. По принципу организации, а для них характерна центральная масса клеток пищеварительной паренхимы (фагоцитобласт), бескишечные турбеллярии напоминают еще фагоцителлообразных прародителей Metazoa.
Низшие Bilateria, в первую очередь плоские черви (тип Plathelminthes), составляют пятый ярус родословного древа животного мира. Билатеральность, централизация нервной системы, выражающаяся в появлении головного мозга, дифференциация осморегуляторной и выделительной системы (протонефридиев), ясная дифференцировка мезодермы во время развития ставят их на более высокую ступень, чем Radiata. К этому же ярусу кроме плоских червей принадлежат типы Namathel-minthes и Nemertini. Одним из очень характерных общих признаков всех перечисленных типов следует считать отсутствие у них вторичной полости тела, что позволяет под названием Acoelomata противопоставлять их всем высшим Bilateria, для которых, напротив, характерно развитие це-лома (Coelomata). Как синоним Acoelomata некоторыми зоологами употребляется название Scolecidae — низшие черви. Все крупные группы сколецид, вероятно, произошли от ресничных червей или от очень близких к ним, не сохранившихся турбелляриеобразных предков.
Верхний ярус филогенетического древа составляют вторичнополост-ные животные — Coelomata. Среди них имеются как вероятно первично несегментированные формы с общим неметамерным целомом (Echiurida, Sipunculida), так и животные, низшие представители которых имеют ясную метамерию наружных органов, но сохраняющие несегментированный целом (тип Mollusca). Последние, вероятно, произошли от примитивных олигомерных кольчатых червей, обладающих уже наружной метамерией, но лишенных еще сегментации вторичной полости тела. Среди современных кольчецов такие близкие к общему предку Coelomata формы есть. Таков, например, Dinophilus (с. 252), не обладающий еще пара-подиями и щетинками, а также кровеносной системой и состоящий из 5 ларвальных сегментов. По плану строения Dinophilus близок к мета-трохофоре; целом у него не подвергается сегментарному расчленению, что характерно также для многих метатрохофор. Большинство Coelomata, однако, составляют сегментированные животные с метамерно расчлененным целомом. Среди последних различают малосегментные формы (типы Tentaculata, Hemichordata) и многосегментные, или полимерные (типы Annelida, Arthropoda, Chordata). Происхождение Coelomata неясно и спорно. Наиболее вероятной кажется нам гипотеза, по которой их предками считаются турбелляриеобразные животные, давшие начало также современным турбелляриям и немертинам.
Основная черта, определяющая более высокую организацию Coelomata,— возникновение вторичной полости тела. Их эволюция сопровождалась развитием кровеносной системы, превращением протонефридиев в метанефридии, формированием целомодуктов. Таким образом, органи-
20* 579
зация животных при переходе к Coelomata подвергалась серьезной перестройке, носившей прогрессивный ароморфный характер.
Следует, однако, заметить, что некоторые из вновь возникших систем органов остаются у высших червей еще слаборазвитыми. Так, в кровеносной системе обычно отсутствует центральный пропульсаторный орган — сердце (лишь у некоторых плеченогих имеется образование, напоминающее сердце). Почти то же можно сказать о дыхательной системе, органы которой очень часто (тип Tentaculata, часть представителей многощетинковых кольчецов Annelida) наряду с дыхательной несут иную функцию (захват пищи, чувствительная), т. е. не строго специализированы.
Типы Arthropoda и Mollusca берут начало от кольчатых червей: первые от полимерных, вторые от олигомерных. Оба типа представлены вторичнополостными животными, у которых заложившиеся на предыдущем этапе филогенетического развития органы достигают значительной степени совершенства. Кровеносная система приобретает сердце, органы дыхания (жабры р?,ков, ктенидии моллюсков, трахеи насекомых) строго специализируются и служат исключительно для газового обмена.
Большой шаг вперед делает нервная система: центральная и периферическая. В центральной нервной системе у многих представителей обоих типов наблюдается, с одной стороны, сильная концентрация ганглиев, а с другой — увеличение относительных размеров мозга. Поражает сильное развитие органов чувств, особенно глаз, которые у головоногих, высших раков и насекомых не уступают в сложности строения органу зрения человека.
Располагая моллюсков и членистоногих в одном горизонтальном ярусе, мы несколько затруднены в оценке того, который из этих типов более высоко организован.
С одной стороны, моллюски имеют более совершенную кровеносную систему: это единственные из беспозвоночных, обнаруживающие дифференциацию сердца на желудочек и предсердие.
С другой стороны, членистоногие снабжены несравненно более совершенными органами движения: богато расчлененные, способные к очень сложным движениям конечности, а кроме того, крылья. Эти свойства, равно как и развитие высшей нервной деятельности (у общественных насекомых в особенности) и воздушных органов дыхания, привели тип Arthropoda в лице высших его представителей, т. е. паукообразных и насекомых, к широкой адаптации к наземному образу жизни.
Кроме позвоночных, мы ни у одной группы животных не встречаем таких многочисленных и разнообразных приспособлений к разным условиям сухопутного существования.
Среди моллюсков выход на сушу наблюдается несравненно реже и не влечет к столь пышному расцвету форм, какой наблюдается у перешедших к наземной жизни представителей типа Arthropoda, а именно у насекомых.
Обсуждая нашу схему родословного древа, мы говорили до сих пор лишь о горизонтальных разделениях — ярусах. Но кроме того, можно провести вертикальную линию, рассекающую Coelomata на две большие группы: первичноротые—Protostomia и вторичноротые — Deutero-stomia. Очевидно, эти филогенетические ветви имели общее начало где-то среди первобытных Coelomata, а затем развивались самостоятельно и параллельно друг другу. Возможно, что некоторые примитивные группы Coelomata, в первую очередь Sipunculida и Tentaculata, в какой-то мере являются промежуточными между первичноротыми и вторичноротыми, и мы лишь условно относим их к Protostomia.
Главные различия между Protostomia и Deuterostomia эмбриологического порядка уже приводились (с. 289). Ход эволюции Protostomia нами только что рассмотрен.
Исследования последних лет показывают, что кроме двух основных эволюционных путей развития Coelomata: первичноротых и вторичноро-тых, есть и другие независимые пути эволюции, которые, возможно, берут начало от неизвестных нам общих предков Coelomata. Такими независимыми линиями развития следует считать щупальцевых (Tenta-culata), щетинкочелюстных (Chaetognatha) и погонофор (Pogonophora) (см. схему с. 578). Последние обладают четырьмя сегментами (а не тремя, как полагали еще недавно), что не позволяет непосредственно включать погонофор во вторичноротых, с которыми они обнаруживают многие черты сходства (в том числе энтероцельный способ образования целома). На организацию погонофор сильный отпечаток наложило обитание в длинных защитных трубках, погруженных нижним концом в ил.
Эволюция Deuterostomia гораздо менее ясна, чем Protostomia. Тип Echinodermata в смысле высоты организации стоит приблизительно уже на уровне высших червей. В самом деле, наличие целома и кровеносной системы уравнивает иглокожих с высшими червями. Дыхательная система иглокожих не строго специализирована, как и у высших червей, ибо кожные жабры их большей частью несут также иные функции: например, через жабры идут процессы выделения (выход амебоцитов) и т. д. Кровеносная система еще не централизована, а нервная находится даже на низшей ступени централизации, чем у кольчатых червей.
Второй, относящийся ко вторичноротым, тип Hemichordata также стоит на низшей ступени развития этой ветви животного царства. Организация обоих типов сводится к трехсегментному строению, столь характерному для диплеврулы, в которой естественно видеть рекапитуляцию общего предка всех вторичноротых. У Hemichordata наблюдается явный переход к полимерному состоянию путем вторичной метамеризации туловищного сегмента.
У вторичноротых, следовательно, также можно различать ларваль-ные и постларвальные сегменты. Однако полимерность у них достигается совершенно иным способом, чем у первичноротых. У Deuterostomia постларвальные сегменты представляют многочисленные метамерные участки сильно вытянутого заднего (третьего ларвального) сегмента, тогда как у Protostomia они всегда образуются на заднем конце тела как добавление к последнему ларвальному сегменту. Таким образом, постларвальные сегменты первичноротых и вторичноротых не гомологичны друг другу.
Наконец, третий тип вторичноротых, а именно Chordata, находится на уровне высших Protostomia и даже поднимается выше. Нервная система хордовых достигает степени дифференциации, неизвестной среди беспозвоночных. То же следует сказать об органах слуха, тогда как глаза хордовых не превышают по сложности строения глаза высших беспозвоночных, а именно головоногих моллюсков. Кровеносная система достигает высшего во всем животном мире развития, хотя у низших представителей хордовых она устроена просто. Выделительная система обнаруживает характер целомодуктов, но чрезвычайно усложнена. Дыхательная система строго специализирована.
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 3178; Нарушение авторских прав?; Мы поможем в написании вашей работы!