КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Колебания в гликолизе
|
|
|
|
Классическим примером колебательной биохимической реакции является гликолиз. В процессе гликолиза осуществляется распад глюкозы и других сахаров, при этом соединения, содержащие шесть молекул углерода, превращаются в трикарбоновые кислоты, включающие три молекулы углерода. За счет избытка свободной энергии в процессе гликолиза на одну молекулу шестиуглеродного сахара образуются две молекулы АТФ. Основную роль в генерации наблюдаемых колебаний концентраций компонентов реакции: фруктозо-6-фосфата, фруктозо-1,6-фосфата и восстановленного НАД (никотинаминадениндинуклеотид) играет ключевой фермент гликолитического пути - фосфофруктокиназа (ФФК).
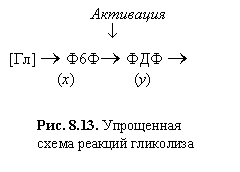
| Рис. 8.13. Упрощенная схема реакций гликолиза |
| Активация ¯ [Гл] ® Ф6Ф® ФДФ ® (x)(y) |
 Упрощенная схема реакций представлена на рис. 8.13:
Упрощенная схема реакций представлена на рис. 8.13:
На схеме [Гл] - глюкоза, Ф6Ф - фруктозо-6-фосфат - субстрат ключевой реакции, ФДФ ‑ продукт этой реакции, который является субстратом в следующей стадии. Обе реакции катализируются ферментами. В безразмерных переменных система описывающих реакции уравнений может быть записана в виде:
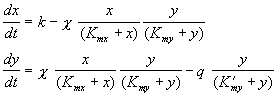
Здесь зависимости скоростей реакций от переменных записаны в форме Михаэлиса-Ментен (Моно), как это было представлено в уравнении (8.10).Если выполняются условия: Kmx>>x, Kmy>>y, можно произвести замену переменных
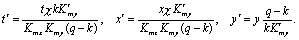
Опустив штрихи у новых переменных, получим систему в безразмерном виде:
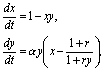 (8.10)
(8.10)
где
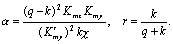
Кинетика изменений переменных и фазовые портреты системы (8.10) при разных значениях параметров представлены на рис. 8.14
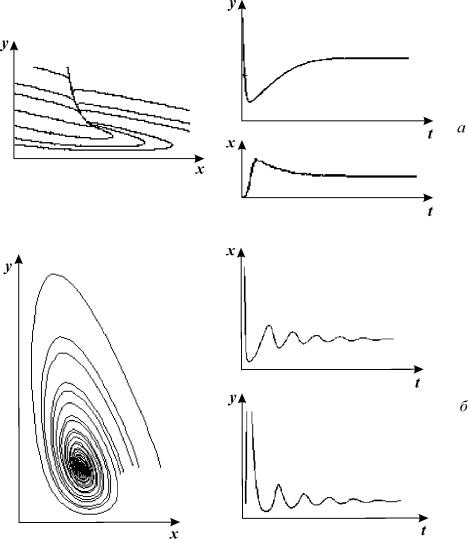
Рис. 8.14. Модель гликолиза (8.10). Кинетика изменений концентраций фруктозо‑6‑фосфата (х) и фруктозодифосфата (у) (справа) и фазовый портрет системы (слева) при разных значениях параметров системы, а ‑ бесколебательный процесс (узел на фазовой плоскости), α = 0.25; r = 1. б – затухающие колебания (устойчивый фокус на фазовой плоскости) α = 4; r = 0.2

Рис. 8.14. Модель гликолиза (8.10). Кинетика изменений концентраций фруктозо‑6‑фосфата (х) и фруктозодифосфата (у) (справа) и фазовый портрет системы (слева) при разных значениях параметров системы, в – колебания с постоянной амплитудой и фазой, близкие к гармоническим (предельный цикл на фазовой плоскости), α = 6; r = 0.2. г – релаксационные колебания с постоянной амплитудой и фазой, (предельный цикл почти треугольной формы на фазовой плоскости) α = 8; r = 0.5
Интересно, что колебательные реакции в системе гликолиза были сначала предсказаны на математической модели (Higgins, 1964), и лишь после этого зарегистрированы экспериментально с помощью метода дифференциальной спектрофотометрии в лаборатории Б. Чанса (1966).
Внутриклеточные колебания кальция.
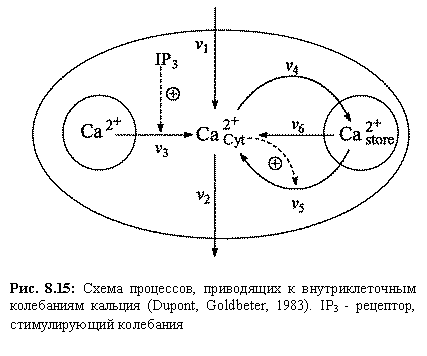
Во многих типах живых клеток наблюдаются колебания внутриклеточной концентрации кальция, период которых может варьировать от 0,5 до 10 мин. Простейшая схема процессов, приводящих к гармонально обусловленным колебаниям кальция, основой которых служит кальций индуцированный выходакальция из клетки, приведена на рис. 8.15. Такие колебания впервые наблюдались Эндо с соавторами (1970) на клетках скелетных мышц, Фабиато (1975) на клетках саркоплазматического ретикулума сердца быка, и позднее — многими другими исследователями.
Схема и модель процессов, предложена и описана в [Dupont and Goldbetter (1989, 1994)]. Рассматриваются приток и отток кальция в клетку через плазматическую мембрану (константы скоростей v 1 и v 2, соответственно); гормонально активируемое освобождение кальция из пула (скорость v 3); активный транспорт цитозольного кальция в пул, (v 4), освобождение кальция из пула, активируемое цитозольным кальцием (v 5); свободный отток кальция из пула в цитозоль (v 6). Модель состоит из двух дифференциальных уравнений
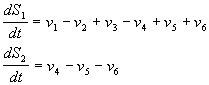 (8.11)
(8.11)
| Рис. 8.15: Схема процессов, приводящих к внутриклеточным колебаниям кальция (Dupont, Goldbeter, 1983). IP3 ‑ рецептор, стимулирующий колебания |
 Здесь S1 ‑ концентрация кальция в цитозоле, S1‑ концентрация кальция в гормонально чувствительном пуле.
Здесь S1 ‑ концентрация кальция в цитозоле, S1‑ концентрация кальция в гормонально чувствительном пуле.
Выражения для величин скоростей были предложены в Simogyi, Stuckin (1991):
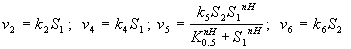 (8.12)
(8.12)
Модель предсказывает колебания концентрации кальция во времени, по форме близкие к экспериментальным (рис. 8.16).
| Рис. 8.16: Модель внутриклеточных колебаний кальция. Кинетика концентрации Ca при разных значениях параметров. a ‑ б ‑ (Dupont, Goldbeter, 1983). |
| a |
| b |
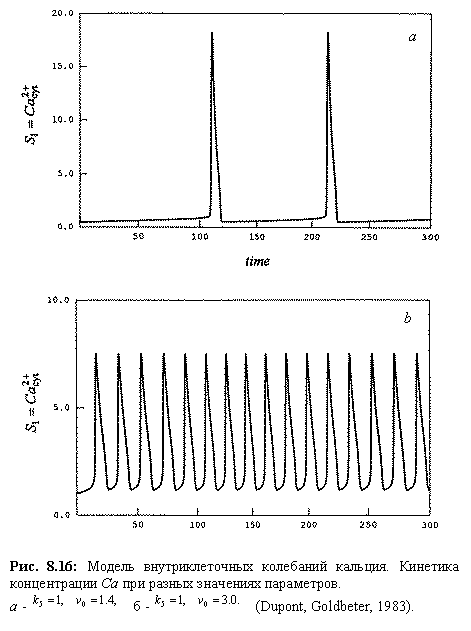
| Рис. 8.17. Схема регуляции клеточного цикла |
| Репликация ДНК |
| Митоз |
| Деградация циклина |
| Деградация циклина |
| G1-циклин |
| Митотический циклин |
 Клеточные циклы.
Клеточные циклы.
В процессе жизненного цикла клетка удваивает свое содержимое и делится на две. В организме млекопитающего для поддержания жизни производятся ежесекундно миллионы новых клеток. Нарушение регуляции пролиферации клеток проявляется как онкологическое заболевание. Этим вызван большой интерес к изучению и моделированию механизмов регуляции клеточного деления.
Схема клеточного цикла изображена на рис. 8.17. Клеточный цикл состоит из двух периодов: митоз (М ‑фаза) включает разделение предварительно удвоенного ядерного материала, деление ядра и деление самой клетки ‑ цитокинез и занимает около часа. Значительно более длительный период между двумя митозами занимает интерфаза, включающая стадию роста G 1, фазу репликации ДНК (S), фазу подготовки к делению G 2. Клеточный цикл (рис.8.17) регулируется генами и белками-ферментами двух основных классов.
| Рис. 8.18. Схема стадий клеточного цикла |
Циклин-зависимые протеин‑киназы (Cdk) индуцируют последовательность процессов путем фосфорилирования отдельных белков. Циклины, которые синтезируются и деградируют при каждом новом цикле деления, связываются с молекулами Cdk и контролируют их способность к фосфорилированию, без циклина Cdk не активны. Количество этих молекул‑регуляторов различно в разного вида клетках.
В делении дрожжевой клетки основные роли играют один Cdk и девять циклинов, которые образуют девять разных циклин‑Cdk комплексов. У гораздо более сложно организованных млекопитающих изучено шесть Cdk и полтора десятка циклинов. Контроль выхода клетки из G 1, и G 2 фаз осуществляют промотор‑фактор S‑ фазы (SPF) и промотор‑фактор M ‑фазы (MPF), представляющие собой гетеродимеры. Cуществует особая контрольная точка клеточного цикла (Start), с которой заканчивается рост (G 1 фаза) и начинается процесс синтеза ДНК.
Простая модель процесса предложена Тайсоном (Tyson, 1995). Постулируется существование фактора транскрипции SBF, который может быть в активной S a и пассивной S i форме. Он переходит в активную форму под действием циклина Cln (N) и Start‑киназы (Cdc28-Cln3) (A) и инактивируется другим веществом (Е). Циклин продуцируется путем активации SBF и деградирует. SBF активируется Chu и Start-киназой и инактивируется фосфатазой. Безразмерная модель процессов имеет вид:
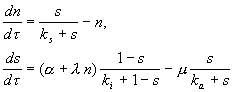 (8.13)
(8.13)
Модель имеет одно или три стационарных решения (два устойчивых) в зависимости от значений параметров, и при увеличении параметра a (в процессе роста клетки) описывает переключение системы из G 1 в S фазу.
Добавление двух уравнений сходного вида позволяет описать также переключение из G 2 в фазу митоза М. Полная модель, учитывающая и другие регуляторные ферменты в фосфорилированной и дефосфорилированной форме содержит 9 нелинейных уравнений (Novak, Tyson 1993) и хорошо описывает кинетику деления ооцитов Xenopus. При соответствующем подборе параметров она применима к описанию деления других типов клеток.
Большое количество работ было посвящено попыткам моделирования периодического воздействия на клеточный цикл с целью оптимизации параметров рентгено‑ радио‑ или хемотерапии при воздействии на клетки онкологических опухолей.
В современной литературе по математической биологии рассмотрены сотни автоколебательных систем на разных уровнях организации живой природы. Несомненно, колебательный характер процессов — эволюционное изобретение природы, и их функциональная роль имеет несколько разных аспектов.
Во-первых колебания позволяют разделить процессы во времени, когда в одном компартменте клетки протекает сразу несколько различных реакций, организуя периоды высокой и низкой активности отдельных метаболитов. Во-вторых, характеристики колебаний, их амплитуда и фаза, несут определенную информацию и могут играть регуляторную роль в каскадах процессов, проходящих на уровне клетки и живого организма. Наконец, колебательные (потенциально или реально) системы служат локальными элементами распределенных активных сред, способных к пространственно‑временной самоорганизации, в том числе к процессам морфогенеза. Эту роль колебательных систем мы подробно рассмотрим во второй части лекций.
Литература
Андронов А.А., Витт А.А., Хайкин С.Э. Теория колебаний. М., Наука, 1981.
Белюстина Л.Н., Кокина Г.А. Качественное исследовани е уравнений фотосинтеза. – В сб. Колебательные процессы в биологических и химических системах. М., Наука, 1967
Гленсдорф П., Пригожин И. Термодинамическая теория структуры, стабильности и флуктуаций. М., 1978
Николис Ж, Пригожин И. Самоорганизация в неравновесных системах. М., Мир., 1979
Чернавский Д.С., Чернавская Н.М. О колебаниях в темновых реакциях фотосинтеза. – В сб. Колебательные процессы в биологических и химических системах. М., Наука, 1967
Dupont G. and Goldbetter A. Theoretical insights into the origin of signal-induced calcium oscillations, in Goldbeter A. (ed). Cell to Cell signalling: From experiments to theoretical models. Acad. Press, London, pp. 461-474, 1989
Dupont G. and Goldbetter A. Oscillations and waves of citosolic calsium: insights from theoretical models. Bioessays, 14, 485-493, 1992
Higgins J.A. A chemical mechanism for oscillations in glicolitic intermediates in yeast cells. Proc. Nat. Acad. Sci. USA, v.51, 1954
Higgins J.A. The theory of oscillating reactions. Ing. Chem. V.59, N5, 1967
Hopf E. Abzweilung einer periodischen Losung von einer stationaren Losung eines Differentialsystems (bifurcation of a periodic solusion from stationary solution of a system of differential equations) Ber. Math-phys. Kl. Sachs. Akad. Wiss. Leipzig 94, 3-22, 1942
Novak B, Tyson J.J. Modeling the cell division cycle: M-phase trigger, oscillation and size control, J. Theor. Biol. 165, 101-104, 1993
Somogyi R, Stuckin J.W. Hormone-induced calcium oscillations in liver cells can be explained by a simple one pool model. J. Biol. Chem.266, 11068-11077, 1991
Prigozhine I.R., Lefebre R. Simmetry breaking instabilities indissipative systems.J. Che. Phys., 48, 1665-1700, 1968
Winfree A.T. The Geometry of Biological Time. Springer, 1980
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 1331; Нарушение авторских прав?; Мы поможем в написании вашей работы!