КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Отдел диатомовые водоросли – Bacillariophyta
|
|
|
|
ЦАРСТВО Heterokontobiontes (Chromista) – Гетероконты
Отдел называется Диатомовые водоросли (от греч. di - два, tome - разрез, рассечение), или Бациллярии (bacillum – палочка). Включает одноклеточные одиночные или колониальные организмы, почти всегда микроскопической величины; формы, видимые простым глазом и достигающие 2–3 мм. Характерно наличие двустворчатого кремнеземного панциря. Известно около 6–10 тыс. видов.
Строение клетки.
Хлоропласты диатомовых водорослей разнообразной формы, обычно пристенные, содержат пигменты – хлорофиллы a и c, каротины, фукоксантины.
У центрических диатомей хлоропласты многочисленные, мелкие, у пеннатных крупные, часто лопастные. Окраска хлоропластов бурая, желтоватая или золотистая. Она обусловлена тем, что зеленые пигменты – хлорофиллы – маскируются добавочными желто-коричневыми ксантофиллами, из которых преобладает фукоксантин. Пиреноидов может быть один или несколько, они выступают за пределы хлоропласта и иногда пронизаны тилакоидами.
В цитоплазме множество капель масла. В виде крупных капель с характерным голубым блеском встречается волютин.
Митохондрииу диатомовых водорослей разнообразной формы (шаровидные, овальные, палочковидные, нитчатые). Аппарат Гольджирасположен рядом с ядром, он состоит из нескольких диктиосом (до 20), которые содержат от 4 до 12 цистерн.
Поверх плазмалеммы у диатомовых формируется особый клеточный покров – панцирь.Он состоит из аморфного кремнезема, сходного по составу с опалом, поэтому диатомовые водоросли часто называют "драгоценными камнями" морей ("опал" в переводе с санскрита означает "драгоценный камень"). Помимо кремнезема в состав панциря входит примесь органических соединений и некоторых металлов. Изнутри и снаружи он покрыт тонким органическим слоем, состоящим из пектиновых веществ. После смерти водоросли содержимое клетки разрушается и исчезает, тогда как кремнеземистый остов оболочки остается без изменений. Он не поддается гниению.
Панцирь состоит из двух половинок: верхней большей – эпитеки и нижней меньшей – гипотеки. Эпитека надевается на гипотеку, как крышка на коробку (рис. 20). В свою очередь эпитека состоит из эпивальвы (верхняя створка) и эпицингулюма (поясковый ободок эпитеки); гипотека состоит из гиповальвы (нижняя створка) и гипоцингулюма (поясковый ободок гипотеки). Два поисковых ободка, накладываясь друг на друга, образуют поясок.У диатомей различают и используют при идентификации две проекции панциря: вид со створки и с пояска. Створка обычно плоская. Ее загнутые край называют загибом створки;он может быть низким или высоким. У некоторых родов (Мелозира, Гиалодискус) створки панциря смыкаются непосредственно краями загибов створок, а поясок образуется во время деления клетки.
Вставочный ободок возникает между краями загиба створки и поясковым ободком (рис. 23). Вставочных ободков может быть много, но самый молодой всегда будет около загиба створки, а наиболее старый около пояска. Значение этих ободков заключается в обеспечении роста панциря и увеличении объема клетки. У ряда видов от внутренних стенок вставочных ободков в полость клетки врастают тонкие кремниевые неполные перегородки – септы. Они всегда имеют одно или несколько отверстий и перегораживают клетку на полуизолированные камеры, что увеличивает поверхность клеток.
Различают два основных типа створок: актиноморфные, через которые можно провести три или более осей симметрии (такие створки характерны для центрических диатомей), а также зигоморфные, через которые можно провести не более двух осей симметрии (такие створки характерны для пеннатных диатомей).

Рис. 23. Диатомовая водоросль Пиннулярия (по:): А – вид со стороны пояска; Б – вид со стороны шва; В – продольный разрез; Г – поперечный разрез; Д – вегетативное размножение; 1 – эпитека, 2 – гипотека, 3 – шов, 4 – узелок, 5 – хроматофор, 6 – пиреноиды, 7 – цитоплазма, 8 – ядро, 9 – вакуоль, 10 – створка, 11 – поясок
Панцирь диатомей пронизан отверстиями-перфорациями, которые служат для сообщения протопласта клетки с внешней средой. Перфорации на створке занимают 10–75 % ее площади. Они могут складываться в ряды, которые видны как штрихи.Штрихи могут быть радиальными, параллельными, конвергентными. Прочность панцирю придают утолщения, выступающие над наружной или внутренней поверхностью створки, называемые ребрами.На поверхности створки часто образуются шипы, щетинки, выступы, шипики, участвующие в образовании колоний.
У некоторых пеннатных диатомей имеется шовная система. Щелевидный шовсостоит из пары продольных щелей (ветвей шва),которыерасположены на створке. Каналовидный шовимеет вид трубки, расположенной в толще створки, гребневидном утолщении ее – киле, или крыловидном выросте, окружающем створку по краю - крыле. Каналовидный шов сообщается с внешней средой с помощью тонкой щели, с внутренней полостью клетки – с помощью отверстий. Швы диатомей обеспечивают сообщение протопласта с внешней средой, а также принимают участие в движении.
Ядрокрупное, содержит 1–8 ядрышек; они исчезают во время митоза.Центриоли отсутствуют. Центром организации микротрубочек являются пластинки (рис. 24). Они расположены на полюсах веретена. Микротрубочки веретена формируются вне ядра, затем проходят в ядро через разрушенные участки его оболочки; ядерная оболочка постепенно исчезает. Таким образом, у диатомовых водорослей митоз открытый. На ранних этапах микротрубочки идут от полюса к полюсу. Кинетохоры хромосом, по-видимому, прикрепляются кполюсным микротрубочкам. В анафазе хромосомы двигаются к полюсам, в поздней анафазе веретено удлиняется. Цитокинез осуществляется за счет образования борозды деления путем впячивания клеточной мембраны от периферии к центру. Цитокинез завершается вместе с формированием оболочек дочерних ядер. Плоскость деления клеток диатомей всегда проходит в плоскости, параллельной створке.
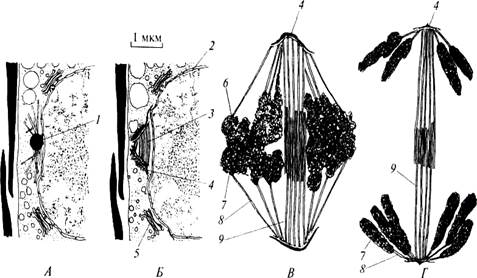
Рис. 24. Митоз у пеннатных диатомей (по: С. Hoek van den et al., 1995): А – интерфаза, почти перед профазой; Б – профаза; В – метафазное веретено деления; Г – анафазное веретено деления; 1 – микротрубочковый центр; 2 – оболочка ядра; 3 – профазное веретено; 4 – полярная пластинка; 5 – аппарат Гольджи; 6 – кинетохор; 7 – хроматида; 8 – хромосомная микротрубочка; 9 – межполюсная микротрубочка
Движение.
Свободноживущие бациллярии передвигаются по субстрату, как бы ползая на стороне створки. Движение происходит равномерно или толчками по направлению продольной оси клетки, притом попеременно, то в одну сторону, то в другую, прямо ей противоположную. Диатомеи, имеющие шов, способны к активному скользящему движению. Это движение осуществляется со скоростью 0,2–25 мкм/с. Относительно механизма их движения выдвинут ряд гипотез. Одна из гипотез связывает движение диатомей с освобождением через шов слизи, в состав которой входят фибриллярные полисахариды. Превращаясь в тяжи, которые выбрасываются вперед по субстрату, они обеспечивают движение диатомей. Считают, что белки кинезин и/или динеин являются той движущей силой, которая приводит к выбрасыванию этих тяжей. Другая гипотеза связывает движение диатомей с трением цитоплазмы, циркулирующей в шве. Трение струящейся цитоплазмы о субстрат развивает моторную силу, перемещающую клетку в направлении, противоположном ее потоку. Третья гипотеза объясняет движение попеременным вбиранием воды и выбрасыванием ее с противоположного конца клетки. Выход потока воды изменяет распределение гидростатического давления в теле водоросли, в силу чего последняя и передвигается в сторону, обратную потоку.
Размножение. У диатомей встречается вегетативное и половое размножение (изогамия, гетерогамия и оогамия).
Половой процесс у центрических и некоторых пеннатных диатомейоогамный.
Яйцеклеток одна или две; они оплодотворяются внутри оогонияили, реже, после их выхода в воду. После оплодотворения формируется диплоидная зигота, которая развивается в растущую ауксоспору.Она одевается оболочкой, постепенно приобретающей характерную для данного вида структуру, превращаясь в вегетативную клетку. У большинства пеннатных диатомей половой процесс изогамный, но у гамет отсутствуют жгутики (рис. 25, А). Перед изогамией происходит мейоз, в результате него формируются 1–2 гаплоидные гаметы. Слияние происходит таким образом, что гамета из одной клетки переползает в другую. Подвижные гаметы можно рассматривать как мужские, а оставшиеся на месте – как женские.
Вегетативное размножение. Чаще размножение диатомовых водорослей происходит вегетативно, путем деления клетки надвое. При делении обе створки раздвигаются до взаимного касания краями, затем клеточное содержимое делится на две симметричные половины в плоскости пояска. Каждая из двух вновь возникших клеточек вырабатывает недостающую ей половинку панциря (створку), вдвигая ее в старую половинку, доставшуюся ей по наследству. У свободноживущих диатомей молодые клетки быстро разъединяются и расходятся, у колониальных же остаются друг подле друга. Так как при дальнейших повторениях процесса деления новые створки всегда вкладываются в старые, и следовательно, меньше их, то после целого ряда делений поколение диатомовых должно значительно измельчать. Клетки, достигнув минимума роста, возвращаются к своей первоначальной величине различными путями. Во-первых, меньшая из образовавшихся клеток может больше не делиться. Во-вторых, у некоторых диатомей пояски панциря более эластичны. В-третьих, возможно раздвигание частей панциря и выравнивание разницы в размерах между эпитекой и гипотекой. В-четвертых, уменьшению размеров клеток у диатомей противопоставляется увеличение их в результате полового процесса, путем образования ауксоспор.
При образовании ауксоспоры происходит процесс копуляции, ведущий к образованию зигоспоры. Две клетки сближаются друг с другом, сбрасывают створки, их протопласты сливаются воедино, окружаются плотной целлюлозной оболочкой, затем увеличиваются в объеме и превращаются в ауксоспору, дающую начало новой особи, заметно превосходящей по величине своих родителей.
Редукционное деление у диатомовых водорослей наблюдается перед образованием гамет, поэтому вегетативные особи представляют собой диплоидные организмы, и жизненный циклдиатомей является диплобионтным с гаметической редукцией.
Покоящиеся стадии. При наступлении неблагоприятных условий диатомовые могут формировать споры и покоящиеся клетки. Эти структуры богаты запасными веществами, которые потребуются при прорастании. Покоящиеся клетки морфологически очень близки к вегетативным клеткам, в то время как панцирь спор становится более толстым, округляется, изменяется его орнаментация. Покоящиеся клетки обычно возникают в условиях среды с низким содержанием растворенного кремния, а споры, наоборот, требуют наличия достаточного количества кремния для построения собственной толстой оболочки. Покоящиеся клетки формируются чаще пресноводными центрическими и пеннатными диатомеями, в то время как споры образуются центрическими морскими диатомовыми. И покоящиеся клетки, и споры могут выживать десятилетиями.
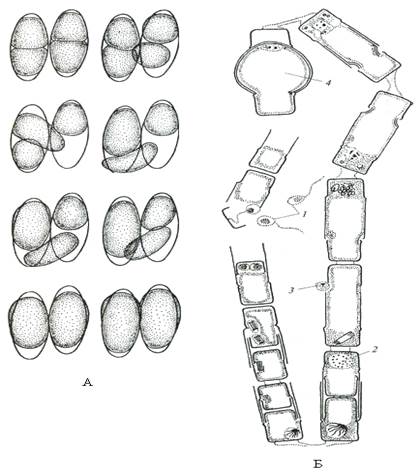
Рис. 25. Схема полового процесса и образование ауксоспор пеннатных диатомовых водорослей (А) – на примере Гомфонемы и центрических диатомовых водорослей (Б) на примере Мелозиры (по: Л. Л. Великанов и др., 1981): 1 – развитие сперматозоидов; 2 – развитие яйцеклетки; 3 – оплодотворение; 4 – образование ауксоспоры
Систематика
Своеобразное движение и кремнеземный панцирь давали повод не раз причислять бациллярий к животным. Теперь общепринято, что диатомеи – водоросли. Большинство исследователей выделяет их в особый отдел. Считается, что к отделу Диатомовые водоросли относится около 6–12 тыс. видов, но некоторые авторы убеждены, что истинное количество видов диатомей может достигать 1 млн. В большинстве систем диатомовые рассматриваются в ранге отдела Bacillariophyta с двумя классами: Центрические – Centrophyceae и Пеннатные – Pennatophyceae.
Класс Пеннатные диатомеи – Pennatophyceae
К пеннатным диатомовым относят обычно подвижные одноклеточные и колониальные представители, через створку которых можно провести одну или две оси симметрии, створки имеют шов. Половой процесс изогамный.
Род Навикула имеет одиночные клетки, обычно подвижные, реже заключенные в студенистые влагалища, простые или разветвленные, внутри которых клетки сохраняют подвижность (рис. 26, В). Створки продольно симметричные, по форме от линейных до эллиптических. Структура створок состоит из поперечных штрихов, расположенных параллельно и радиально. Два крупных пластинчатых хроматофора расположены вдоль клетки и прилегают к ее пояску. Виды широко распространены в пресных и солоноватых водоемах, реже встречаются в морях. В основном бентические и, редко, планктонные формы.
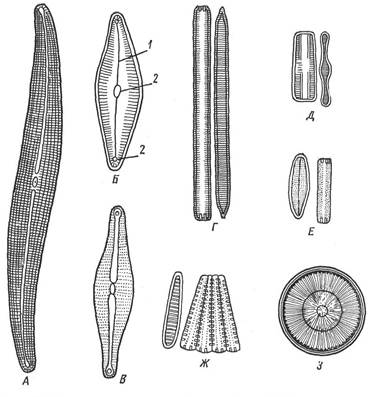
Рис. 26. Диатомовые водоросли (по:): А – Плеуросигма; Б – Цимбелла; В – Навикула; Г – Синедра; Д – Табеллярия; Е – Диатома; Ж – Меридион; З – Циклотелла: 1 – шов, 2 – узелок
Род Диатома – клетки соединены в лентовидные и зигзаговидные колонии, иногда в звездчатые (рис 26, Е). Панцирь с пояска линейный, с прямыми углами, вставочные ободки иногда имеются, септы отсутствуют. Створки от эллиптических до линейных, структура створок из поперечных грубых ребер и нежных поперечных штрихов. Осевое поле нитевидное, едва заметное. Римопортулы располагаются на концах створки. Хроматофоры мелкие, зернистые, многочисленные. Виды пресноводные, преимущественно бентические.
Род Табеллярия – клетки прямоугольные с пояска, эллиптические до удлиненно линейных (рис. 26, Д, рис. 27, Г). Имеются вставочные ободки с септами. Шов отсутствует. Пресноводные эпифиты.
Род Ницшия –клетки от палочковидных до эллиптических, прямые, реже изогнутые, одиночные, очень редко соединенные в нитевидные или разветвленные колонии. Створки линейные, реже ланцетные и эллиптические, с килями и поперечными штрихами. В киле находится каналовидный шов. Один пластинчатый хроматофор расположен вдоль клетки по ее диагонали или прилегает к одной из поясковых сторон. Морские, солоноватоводные и пресноводные, чаще бентические виды (приложение, 3Б).
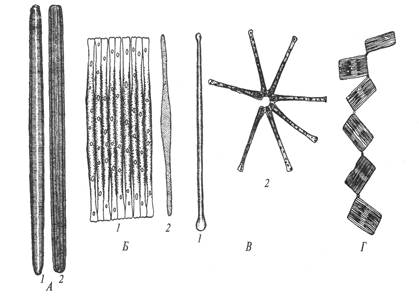
Рис. 27. Бесшовные диатомовые водоросли: А – Синедра; Б – Фрагилярия; В – Астерионелла; Г – Табеллярия: 1 – панцирь со створки, 2 – панцирь с пояска
Класс Центрические диатомеи – Centrophyceae
Центрические диатомеи – одноклеточные и колониальные формы, через створку которых можно провести три или более осей симметрии, у которых отсутствует активная подвижность, не имеется шва на панцире и наблюдается оогамный половой процесс. Виды центрических диатомей очень широко представлены в планктоне морей и океанов как одни из главных продуцентов органических веществ.
Род Косцинодискус –клетки дисковидные, реже линзовиднын, всегда одиночные. Панцирь обычно грубый, часто со вставочными ободками (рис. 28, Б). Створки круглые, плоские, выпуклые, иногда волнистые. По
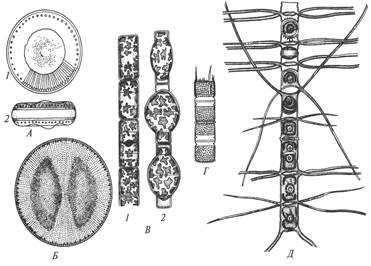
Рис. 28. Центрические диатомовые водоросли: А – Циклотелла; Б – Косцинодискус; В – Мелозира; Г – Аулакозира; Д – Хетоцерос с пояска, часть цепочки: 1 – вид со створки, 2 – вид с пояска
краю створки часто бывает кольцо мелких шипиков, расположенных под определенным углом. Хроматофоры – многочисленные пластинки, расположенные по всей поверхности клетки. Ядро центральное, прилегает к одной из створок, реже подвешено на цитоплазматических тяжах в центре клетки. Виды морские, преимущественно планктонные. Нередко встречаются в большом количестве в опресненных морских участках и во внутренних солоноватых морях.
Род Ризосоления имеет клетки цилиндрической формы. Клетки часто сильно вытянутые в высоту, в виде длинной палочки, прямые, редко слегка изогнутые, одиночные, реже соединенные в нитевидные колонии. Панцирь в поперечном сечении от круглого до эллиптического, очень тонкий. Имеются многочисленные вставочные ободки, кольцевидные, трапециевидные, ромбические и чешуевидные. Створки в виде колпачка с вытянутой верхушкой, конец которой продолжается в виде шипа, щетинки или выроста. Хроматофоры многочисленные, зернистые или дисковидные, часто скапливаются в центре клетки вокруг ядра. Виды морские, планктонные (приложение, 3Б).
Род Хетоцерос – клетки низкоцилиндрические, соединенные в цепочки, реже одиночные (рис. 28, Д). Створки эллиптические, плоские, выпуклые или вогнутые. От полюсов створки отходит по одной тонкой щетинке, с помощью которых клетки соединяются в колонии. При этом щетинки створок смежных клеток в цепочке соприкасаются одна с другой, изгибаются и скрещиваются, довольно часто срастаясь в месте соприкосновения. Конечные щетинки цепочки обычно отличаются от прочих длиной, толщиной и направлением. Отверстия между смежными створками в цепочке называются окнами, очертания которых могут быть самыми различными. Родвключает большое количество исключительно планктонных видов, которые составляют основную массу прибрежного фитопланктона морей и океанов.
Экология и значение
Виды диатомовых водорослей широко распространены в природе, они встречаются во всевозможных биотопах. Диатомовые водоросли обитают в морских, солоноватых и различных пресных водоемах: как в стоячих (озерах, прудах, болотах), так и в проточных (реках, ручьях, оросительных каналах). Они распространены в почве, их выделяют из образцов воздуха, они образуют богатые сообщества во льдах Арктики и Антарктики. Диатомовые водоросли в водных экосистемах доминируют над другими микроскопическими водорослями круглый год. Они обильны как в планктоне, так и в перифитоне и бентосе. В планктоне морей и океанов преобладают центрические формы диатомей, хотя к ним примешиваются и некоторые пеннатные формы. В планктоне пресных водоемов, наоборот, преобладают пеннатные диатомеи. Бентосные (донные) сообщества также отличаются большим разнообразием и количеством диатомовых, которые обычно обитают на глубине не более 50 м. Бентосные бациллярии ползают по субстрату или прикрепляются с помощью слизистых ножек, трубок, подушечек.
Во множестве появляются диатомеи в стоячих водах (прудах, лужах, канавах), образуя на дне и у берега желто-бурые слизистые пленки. Их находят в минеральных источниках, в водопроводах, даже в графинах с водой, на поверхности влажных скал, камней, дерева, земли (например, в горшках с растениями) и т. д. Некоторые виды эпифитных диатомей прикрепляются к морским и пресноводным водорослям, иногда в таком множестве, что водоросли становятся просто неузнаваемыми. После высыхания водоемов диатомовые могут быть подняты вместе с пылью ветром и перенесены на большие пространства. Многие виды настоящие космополиты и встречаются повсюду. Видовой состав бациллярий в водоемах определяется комплексом абиотических факторов, из которых большое значение в первую очередь имеет соленость воды. Виды родов Ризосоления, Скелетонема, Хетоцерос, Биддульфия, Шизонема, Стриателла являются типичными морскими. Морские формы обычно отличаются бóльшими размерами и более крепкими оболочками. В пресных водах живет большинство представителей родов Цимбелла, Фрагилярия, Навикула, Гомфонема. Не менее важным фактором для развития диатомей являются температура, степень освещенности и качество света. Диатомовые водоросли вегетируют в диапазоне 0–700С, но в состоянии покоя способны переносить более низкие или более высокие температуры.
Наиболее богаты по качественному и количественному составу диатомей сообщества обрастания и эпибионтов. Диатомеи занимают господствующее положение среди эпифитов высших растений и макроскопических водорослей в пресных водоемах и морях. Обрастанию диатомовыми водорослями могут подвергаться многие морские животные: от ракообразных до китов. Такие водоросли называются эпизоитами. Среди диатомей имеются и эндосимбионты, которые обитают в бурых водорослях, фораминиферах и др.
Бациллярии имеют огромное значение в природе. Составляя значительную массу фитопланктона, диатомеи являются началом пищевой цепи. Их поедают как зоопланктонные организмы, так и молодь и взрослые рыбы. Давая примерно около четверти органики всей планеты, они являются наиболее важными продуцентами органического вещества в Мировом океане. Бациллярии играют основную роль в круговороте кремния, ежегодно поглощая из Мирового океана около 3 млрд. тонн.
Билинский (г. Билин в Богемии) полировальный сланец, залежи которого, толщиной в 0.6–5 метров, раньше других стали известными, весь состоит из панцирей диатомей, причем главная масса слагается одним только видом, еще и теперь живущим – Melosira distans. Большая залежь люнебургских верещатников достигает до 13 м толщины; она состоит преимущественно из Synedra ulna. Залежь около Берлина достигает 30 м; на ней стоит часть самого города. Подобная залежь находится около Кенигсберга. Все эти залежи недавнего происхождения. Но существуют и гораздо более древние, относящиеся к третичной эпохе. На такой залежи расположен, например, город Ричмонд в Соединенных Штатах Америки. Диатомеи найдены также в янтаре (третичное образование). В России залежи сланцев найдены в Ульяновской, Пензенской области и в Сибири.
Важнейшая роль принадлежит диатомеям в осадкообразовании на дне океана. Панцири планктонных диатомей, отлагавшиеся после отмирания водорослей на дне морских и пресноводных бассейнов, образовали там мощные отложения диатомита (горная мука) – масса белого или светло-серого цвета, легкая, пористая и твердая. Диатомит на 50–80% состоит из панцирей диатомей.
Толщина диатомитового слоя в некоторых местах достигает нескольких сотен метров. Крупные залежи диатомита в России встречаются в Тюмени, Поволжье, Приморском крае и в ряде других мест. Только на территории Тюменской области открытые в прошлом веке залежи диатомитов составляют до 500 × 1012 м3. Считают, что это открытие можно поставить в один ряд с обнаружением за Уралом запасов нефти и газа, так как диатомит – сырье многоцелевого назначения. Он служит источником около 100 разнообразных продуктов, находя применение как материал для получения оптического стекловолокна, жидкого стекла, как фильтрующее вещество в различных отраслях промышленности, как полировочный и шлифовальный материал. Его используют и в качестве строительного материала, а также для изготовления динамита.
Горная мука служит хорошим полировочным средством (трепел), из нее приготовляют легкие кирпичи (кирпичи Фаброни), а иногда и стекло; она идет далее для изготовления динамита, для чего ее смешивают с нитроглицерином. Горная мука в нужде употреблялась в пищу у лапландцев, тунгусов и других полудиких племен (съедобная земля). Горную муку либо прямо ели, либо смешивали с обычной мукой и пекли хлеб. Питательность горной муки, по всей вероятности, обусловлена теми остатками органических веществ, которые еще сохранились в отмерших водорослях.
Панцири диатомей длительное время сохраняются в ископаемом состоянии, поэтому их используют для определения происхождения и возраста различных осадочных пород. Особое значение имеют диатомовые водоросли в экологическом мониторинге, поскольку служат хорошими индикаторами органического загрязнения водной среды.
Однако массовое развитие некоторых видов диатомовых водорослей может иметь и отрицательное значение. У ряда диатомей обнаружены токсины. Так, представители родов Псевдонитшия и Нитшия образуют домоевую кислоту, которая вызывает амнезическое отравление у людей и животных. Эта кислота растворима в воде и нерастворима в органических растворителях. Впервые домоевая кислота была выделена из макроскопической красной водоросли Хондрии(по-японски "домой" – отсюда и название кислоты). В 1987 г. в Канаде было зарегистрировано более 100 случаев отравления людей этим токсином после употребления мидий, четверо из пострадавших умерли. Симптомами отравления являются, в легком случае, тошнота, рвота, диарея; в тяжелом случае исчезает болевая чувствительность, появляются галлюцинации, происходит потеря кратковременной памяти. Диатомовые водоросли могут засорять речные устья и гавани, нередко вызывают "цветение" воды и являются причиной появления в ней неприятных запахов. Забивая жабры животных, диатомовые водоросли вызывают их гибель.
Контрольные вопросы
1. Назовите общие характерные признаки диатомовых водорослей
2. Строение панциря диатомовых водорослей.
3. Какие пигменты и типы питания известны у бациллярий?
4. Как размножаются диатомовые водоросли? Жизненный цикл диатомовых водорослей.
5. Дайте характеристику диатомовых водорослей класса пеннатные.
6. Дайте характеристику диатомовых водорослей класса центрические.
7. Назовите типичных представителей диатомей класса пеннатные и центрические.
8. В каких местообитаниях встречаются диатомовые?
9. Значение диатомовых водорослей для природных экосистем.
10. Хозяйственное значение диатомовых водорослей.
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 9890; Нарушение авторских прав?; Мы поможем в написании вашей работы!