КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Обязательная самоподготовка
|
|
|
|
Основная литература
1. Догель В.А. С. 92 – 113, 201 – 202.
2. Шарова И.Х. С. 99 – 120 (Mesozoa – по Догелю).
Дополнительная литература
3. Буруковский Р.Н. С.107 – 168.
4. Беклемишев К.В. Курс лекций. С. 37 – 54.
5. Беклемишев К.В. Метод. указ. С. 19 – 24.
При изучении учебного материала следует проанализировать гипотезы возникновения многоклеточности. Первая группа гипотез колониального происхождения многоклеточности (гетероклеточности) является исходной при формировании современных подходов к решению этой проблемы (Э. Геккель, 1874; О. Бючли, 1884; И.И. Мечников, 1882; А.А. Захваткин, 1949; А.В. Иванов, 1967; В.Н. Беклемишев, 1964; Н.А. Ливанов, 1955; Л.Н. Серавин, 1994 – 2005. и др.). Все многоклеточные животные обязательно имеют оогамный ядерный цикл с гаметической редукцией, т.е. их жизненный цикл состоит из:
1) прогамной части (образование гамет);
2) сингамной - слияние гамет и образование зиготы;
3) палинтомического дробления – метагамной части цикла;
4) монотомической фазы роста (вегетативной части цикла).
Умногоклеточных животных в жизненном цикле нет изогамии, анизогамии и зиготической или промежуточной редукции.
Гипотеза гастреи (Геккель,1874): инвагинация (впячивание) бластулообразного (шаровидного) организма; возникновение первичной кишечной полости (гастреи) с ротовой порой. Однако:
а) инвагинационное образование двухслойности встречается только у Deuterostomia (вторичноротых) - иглокожие, хордовые;
б) гастрея в качестве типичной личиночной стадии у ныне живущих многоклеточных не отмечена; гастрее соответствуют только ныне живущие стрекающие кишечнополостные животные;
в) инвагинация возможна если бластула заполнена первичнополостной жидкостью;
г) предковая форма полой бластулы (целобластулы) не могла существовать миллионы лет поскольку у такой бластулы не было фагоцитирующих клеток.
Гипотеза фагоцителлы (Мечников, 1886). Гипотетические предковые организмы с мерцательным покровом, клетки которого обладали способностью к фагоцитозу (паренхимулы = фагоцителлы) – исходная форма многоклеточных. Это моноаксонные (одноосные), гетерополярные (аборальный и оральный полюсы), радиальносимметричные (с бесконечно большим порядком радиальной симметрии) бионты, которые эволюционировали параллельно с разной «скоростью».
Фагоцителла (паренхимелла) состоит (подобно личинке современных низших многоклеточных – паренхимуле) из слоя поверхностных клеток – эктодермы, или кинобласта (функции отграничения, внешнего обмена и движения), и внутренней клеточной массы – паренхимы, или фагоцитобласта (функция внутреннего обмена, внутриклеточного пищеварения). Из кинобласта и фагоцитобласта в ходе эволюции возникло всё многообразие форм тканей многоклеточных животных организмов.
Способы формирования фагоцитобласта:
а) фагоцитирующие клетки уходят во внутрь мультиполярно образуя паренхимулу. Сейчас на этом уровне развития находятся паренхимулы губок, планулы книдарий (стрекающих);
б) униполярная иммиграция – фагоцитирующие клетки более крупные, расположены на оральном полюсе, на котором и уходят (иммигрируют) под кинобласт, образуя паренхиму личинки. При униполярном типе развития возможны: дифференцировка фагоцитобласта на центральный фагоцитобласт (энтодерма) и периферический фагоцитобласт (третий зародышевый листок - мезодерма) и телобластический тип закладки мезодермы – третьего зародышевого листка и вторичной полости тела целома;
в) инвагинация (впячивание) развивается из униполярного типа. Бластула крупная, с протоцелем (первичная полость тела) с полостной жидкостью, в которой накапливаются метаболиты покровных клеток - сигнальные молекулы, обеспечивающие единовременное перемещение всех фагоцитирующих клеток орального полюса – впячивание. Следующим шагом при инвагинационном типе развития является энтероцельное образование мезодермы и целома: выпячивание карманов в энтодерме и «отшнуровка» этих карманов. Эпителий карманов – это мезодерма, а полость – это целом (вторичная полость тела). При энтероцельном типе развития возникает билатеральная симметрия (двусторонняя симметрия зеркального отражения) и происходит образование целома (диплеврула иглокожих, онтогенез хордовых).
В жизненных циклах современных беспозвоночных, стоящих у истоков филогенетического древа многоклеточных, имеются фагоцителлоподобные стадии трех структурных уровней (рис.4), соответствующие предковым формам:
фагоцителла первая (Ф-1) с одним эпителием (кинобласт) – паренхимула губок, из ныне живущих – трихоплакс, бластулы многоклеточных;
фагоцителла вторая (Ф-2) – с двумя эпителиями (кинобласт и фогоцитобласт), изначально радиально симметричной со склонностью к билатеральной симметрии – амфибластула высших губок (у которых есть тенденция к инвагинации), аскон губок, все стрекающие кишечнополостные, гаструла многоклеточных;
фагоцителла третья (Ф-3) – с тремя эпителиями (кинобласт, центральный и периферический фагоцитобласты), т.е. с экто-, энто- и мезодермой. Ей соответствуют сикон и лейкон губок, диплеврула (личинка иглокожих).
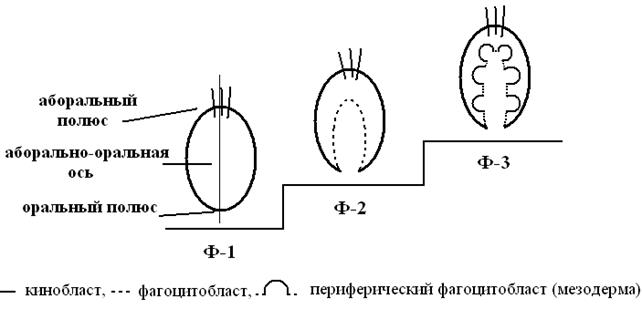
Рис.4. Структура фагоцителл
У потомков Ф-1 и Ф-2 широко варьируют типы яйцеклеток, характер дробления, способы формирования паренхиматозного или эпителизованного фагоцитобласта. У потомков Ф-3 – дробление радиальное, гаструла инвагинационная, мезодерма, формирующая вторичную полость тела (целом), энтероцельная. Все фагоцителлы могли переходить к обитанию на субстрате тремя путями: оседая на аборальный, оральный полюс или «ложась» на одну из антимер (рис.5).

Рис.5. Способы оседания фагоцителл
Сочетания вариантов (типов яйцеклеток, способов дробления яйцеклеток, типов гаструляции, образования третьего зародышевого листка, вторичной полости тела (целома), способа оседания на субстрат) обеспечивают морфологическое разнообразие в эволюции многоклеточных животных.
У свободноживущих животных все стадии перехода от одноклеточных к многоклеточным не прослеживаются. У многоклеточных (гетероклеточных) животных стадии перехода выявляются только на отдельных этапах онтогенеза.
Втораягруппа – гипотезы происхождения многоклеточных от полиэнергидных простейших (целлюляризация). Пути образования многоклеточного животного организма из одноклеточных эукариот разделением исходного многоядерного одноклеточного организма на отдельные клетки. И. Хаджа (1963) считал предками Metazoa инфузорий, а первыми многоклеточным – ресничных червей (планарий). Эти гипотезы не имеют достаточного подтверждения в жизненных циклах ныне живущих беспозвоночных: ни у кого из них целлюляризация не описана. Этот путь не подтверждается и современными ультраструктурными данными.
Третья группа гипотез, сторонники которых считают, что многоклеточные произошли от тех видов амебоидных организмов, которые способны, сползаясь вместе, формировать единый сложный организм.
Наиболее вероятно существование параллельных подходов к многоклеточности у животных, которые могли возникнуть непосредственно в рамках Protista (первичных Sarcomastigophora) и обладают принципиально различными чертами организации. Это: Protozoa, Porifera, Mesozoa (Orthonectida, Dicyemida), Placozoa, Coelenterata (Cnidaria и Ctenophora), Plathelminthes.
У одноклеточных Labirinthomorpha, Ascetpspora и Myxozoa многоклеточность одной из стадий жизненного цикла сочетается с саркодовым типом организации. Orthonectida и Dicyemida (Mesozoa) имеют в жизненном цикле многоядерный плазмодий (саркодовый тип организации), в котором формируются половозрелые, свободноживущие особи с мерцательным эпителием (монадный тип организации). Дальнейшее развитие способа перехода к многоклеточности через саркодовый тип организации не подтверждается жизненными циклами ныне живущих беспозвоночных, у которых хорошо выражена тенденция к построению многоклеточного тела или личиночной стадии с мерцательными покровами: Plaсozoa, Spongia, Coelenterata (Cnidaria и Acnidaria), Plathelminthes.
Среди одноклеточных Саркодовые отличаются относительно низким структурным уровнем, незначительной вариабельностью признаков и жизненных циклов, отсутствием склонности к колониальности. Ciliophora, среди Protozoa, обладают самым высоким структурным уровнем, но у них практически нет вариабельности жизненных циклов и очень слаба тенденция перехода к колониальности. Среди гетеротрофных жгутиконосцев (Protozoa) многоклеточность в жизненном цикле не отмечена, колонии есть только у воротничковых (Choanoflagellata). Однако Phytomastigina представлены как одиночными так и колониальными формами с видоспецифичными жизненными циклами и количеством клеток в колонии. Для Mastigophora характерны максимально широкая вариабельность жизненных циклов, в т.ч. ядерных, при более высоком, по сравнению с саркодовыми, структурном уровне, что обеспечивает возможность эволюционирования к многоклеточным формам.
Таким образом, в животном мире доминирует способ возникновения многоклеточности от первичных Sarcomastigophora через колониальность и монадный тип организации (первая группа гипотез).
Среди многоклеточных животных существуют пять типов с «отрицательными признаками»: у них нет дефинитивных (окончательно установленных) фагоцито- и кинобласта; нет рта, кишечника; никогда не было мускулатуры инервного аппарата.
Надраздел Prometazoa (Прометазои)
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 601; Нарушение авторских прав?; Мы поможем в написании вашей работы!