КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Часть III. Историческое развитие (филогенез) организмов 5 страница
|
|
|
|
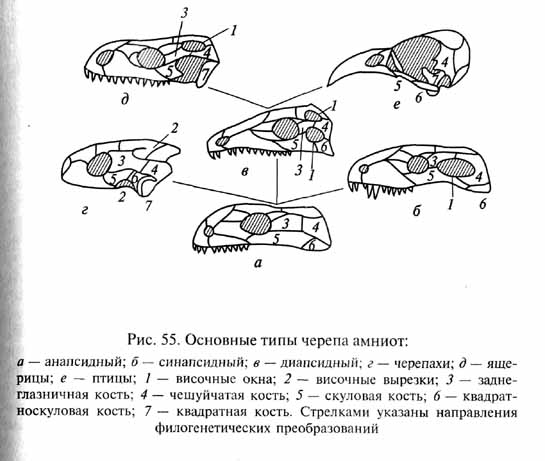
Самые крупные хищники пермского времени принадлежали к подклассу зверообразных, или синапсидных рептилий (Theromorpha, или Synapsida). У этих пресмыкающихся в височной области черепа для его облегчения и предоставления необходимого пространства для сокращения мощных челюстных мышц возникла одна пара широких отверстий, или височных окон, пронизавших наружный черепной панцирь и ограниченных по бокам скуловыми дугами (рис. 55). Такой тип черепа, называемый синап-сидным, дал одно из названий подкласса.
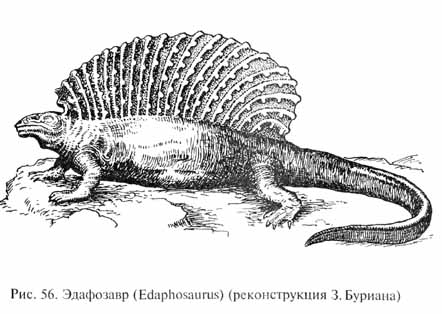
Самыми древними зверообразными рептилиями были пеликозавры (Pelycosauria), появившиеся уже в позднем карбоне. Их расцвет приходится на раннепермское время, когда многие их представители достигали крупных размеров, до 2 м. Некоторые пеликозавры (диметродон - Dimetrodon, эдафозавр - Edaphosaurus и др.) имели странное приспособление: гипертрофированные остистые отростки спинных позвонков поднимались над позвоночником почти на 1 м, а у эдафозавра (рис. 56) на этих отростках имелись еще и короткие боковые выросты. Вероятно, между ними была натянута складка кожи, образуя огромный продольный гребень ("спинной парус"). Может быть, эта складка играла роль в терморегуляции, увеличивая поверхность тела, что могло в одних ситуациях усиливать теплоотдачу, в других же - содействовать повышению температура тела при обогревании в солнечных лучах (повышение температуры тела путем инсоляции, называемое гелиотермией, присуще всем современным рептилиям).
Большинство пеликозавров вымерло уже в середине пермского периода, вероятно не выдержав конкуренции с представителями возникшей от пеликозавров более прогрессивной группы зверообразных рептилий - терапсид (Therapsida). В позднепермской эпохе терапсиды стали доминирующей группой пресмыкающихся. Этому способствовал целый ряд присущих им прогрессивных особенностей, в частности усовершенствование наземного передвижения. У большинства рептилий сохранилось примитивное положение конечностей, при котором плечо и бедро располагаются в целом горизонтально, локоть и колено направлены вбок, и тело животного удерживается на весу между конечностями усилиями мышц (отсюда характерное для рептилий "пресмыкание", когда брюхо при движении лишь незначительно приподнимается над землей). У высших терапсид локтевой сустав повернулся назад, коленный - вперед, бедро и плечо (и конечности в целом) оказались расположенными под телом, высоко приподнятым над субстратом; для поддержки тела при этом не требуется столь значительной работы мышц, как у типичных пресмыкающихся.
У многих терапсид произошла дифференциация зубной системы на передние острые "резцы", крупные кинжалообразные "клыки" и заклыковые щечные зубы, которые у высших представителей этой группы (териодонтов, или "зверозубых", - Theriodontia) приобрели усложненную коронку с тремя бугорками, что позволяло более эффективно пережевывать пищу. Пережевывание пиши в ротовой полости требовало задержки дыхания, в связи с чем у терапсид развивается вторичное нёбо, разделившее примитивную ротовую полость на два этажа: верхний (носовой ход) и нижний (собственно ротовая полость).
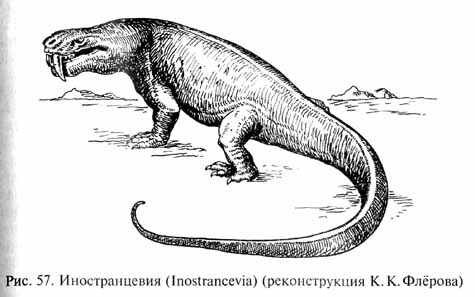
Терапсиды были очень разнообразны: среди них были и хищники различных размеров, вплоть до трехметровых иностранцевий (Inostrancevia, рис. 57) с клыками более 10 см длиной (своего рода "саблезубых тигров" пермского периода), и растительноядные животные, иногда имевшие причудливое строение. Для некоторых растительноядных дейноцефалов (Deinocephalia, их название буквально означает "страшноголовые") характерны большие костные бугры или шишки на темени и щечной области черепа, придававшие голове этих животных устрашающий вид (скорее всего, это были защитные приспособления). Были и хищные дейноцефалы, например приземистый и длинный титанофонеус - Titanophoneus, большие лапы которого, по мнению Ю.А.Орлова, были снабжены плавательными перепонками, а сам этот хищник вел полуводный образ жизни.
В позднепермское время были широко распространены дици-нодонты (Dicynodontia), утратившие все зубы, кроме пары огромных верхних клыков, сохранявшихся у некоторых видов лишь у самцов. Беззубые челюсти были покрыты роговым клювом. Нелегко представить себе способ питания этих странных животных, среди которых были и сравнительно мелкие, и очень крупные, размеров носорога, виды. Предполагали, что дицинодонты питались падалью или поедали питательную сердцевину стволов саговниковых растений, раскалывая или разрывая их своими клыками, или выкапывали корневища хвощеобразных растений, в изобилии произраставших по берегам водоемов.
Местонахождения с ископаемыми остатками целых комплексов видов позднепермских рептилий (котилозавров, дицинодонтов, дейноцефалов, териодонтов) с довольно сходным общим обликом фауны известны в Южной Африке и Восточной Европе. Замечательные местонахождения позднепермских тетрапод были обнаружены на территории России. На берегу Северной Двины, в районе г.Котласа, В.П.Амалицким в 1897г. были найдены остатки крупных парейазавров, дицинодонтов, хищных териодонтов (среди них были огромные иностранцевии), батрахозавров и лабиринтодонтов. Еще одна интересная позднепермская фауна была раскопана в 1957-1960 гг. в Очерском районе Пермской области. В состав Очерской фауны, исследованной П.К.Чудиновым, входили хищные и травоядные дейноцефалы (среди последних огромные рогатые эстемменозухи, Estemmenosuchus), хищные териодонты, терапсиды, близкие к предкам дицинодонтов, лабиринтодонты. Костеносные породы этих местонахождений сформировались из глинистых и песчаных осадков речных дельт1.
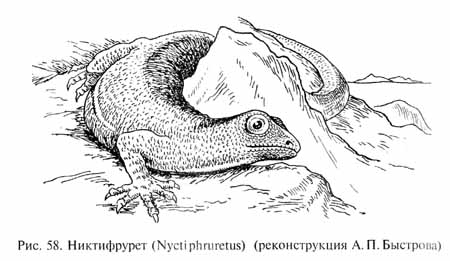
В некоторых филогенетических стволах рептилий продолжалось совершенствование приспособлений к насекомоядности, которая была, вероятно, самой примитивной формой питания пресмыкающихся. Это направление эволюционных преобразований привело к появлению небольших и проворных ящерицеобразных животных, причем сходный тип строения сформировался параллельно в нескольких независимых филетических линиях. Одна из них сохранила общий уровень организации котилозавров, со сплошным покровом накладных костей в височной области черепа. Это были проколофоны (Procolophonia). Остатки примитивных проколо-фонов, никтифрурета (Nyctiphruretus, рис.58) и никтеролетера (Nycteroleter) - легких большеглазых животных с удлиненным телом и хвостом общей длиной до 30-40 см были найдены в верхнепермских отложениях на берегах р. Мезени.
В других группах насекомоядных рептилий произошли некоторые прогрессивные изменения организации, в частности усовершенствование строения черепа. Эти животные, давшие начало двум центральным подклассам рептилий: лепидозаврам (Lepidosauria) и архозаврам (Archosauria) - приобрели удлиненные челюсти, удобные для схватывания мелкой подвижной добычи, и две пары височных окон, расположенных друг над другом и разделенных тонким костным мостиком - верхней височной дугой. Так возник диапсидный тип черепа (см. рис. 55, в), легкая и ажурная конструкция которого позволила усовершенствовать так называемый кинетизм - подвижность верхней челюсти и некоторых связанных с ней костей, унаследованную наземными позвоночными от кистеперых рыб (рис. 59).
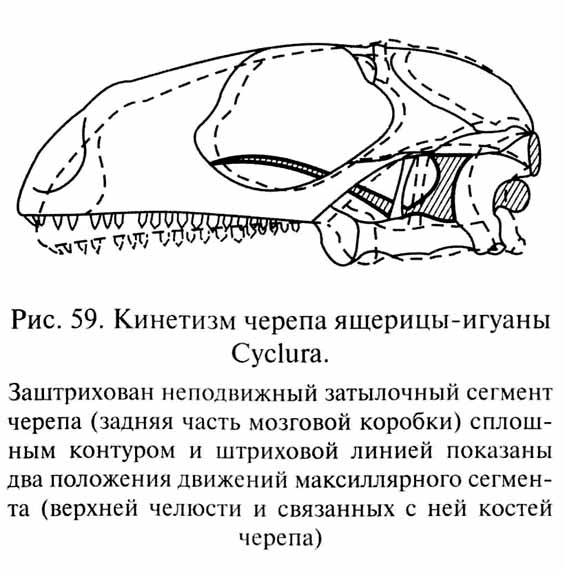
Примитивные лепидозавры (миллерозавры - Millerosauria), у которых лишь началось развитие височных окон и усиление кинетизма черепа, известны из верхнепермских отложений Южной Африки. В породах примерно того же возраста сохранились остатки первых рептилий, имевших настоящий диапсидный череп - эозухий (Eosuchia), наиболее известным представителем которых была янгина (Youngina). Отдаленным потомком эозухий является дожившая до нашего времени новозеландская гаттерия (Sphenodon), которая принадлежит к отряду клювоголовых (Rhynchocephalia), появившихся в раннем триасе. От эозухий или от миллерозавров возникли настоящие ящерицы (Lacertilia), у которых в связи с усилением подвижности верхней челюсти (см. рис. 59) редуцировалась нижняя височная дуга и череп приобрел легкую ажурную конструкцию. Ящерицы достоверно известны начиная со среднего триаса.
Вероятно, эозухий были также предками высших рептилий, принадлежащих к подклассу архозавров, которые оставили самые яркие страницы в палеонтологической истории мезозоя. Д.Уотсон высказал предположение, что обособление предков архозавров от ранних насекомоядных лепидозавров было связано с переходом первых к питанию позвоночными. Этим объясняется более Массивная конструкция черепа архозавров и развитие у них более крупных зубов, сидящих в отдельных ячейках (текодонтность). Для архозавров характерно также более выпрямленное положение конечностей, чем у лепидозавров.
Наиболее ранним известным архозавром является позднепермский проторозавр (Protorosaurus), пропорциями тела и размерами (1-2 м) напоминавший современных ящериц-варанов.
ГЛАВА 3. МЕЗОЗОЙСКАЯ ЭРА - ВЕК РЕПТИЛИЙ
После завершения в середине пермского периода оледенения Гондваны началось потепление, которое продолжалось на протяжении мезозойской эры, в целом более однообразной в климатическом отношении, чем другие эры фанерозоя. В мезозое господствовали теплые климаты с относительно слабо выраженной климатической зональностью. Следов оледенения какого-либо континента в мезозое не обнаружено. Южный полюс оставался в океане, а Северный перемещался от восточной оконечности Сибири к Аляске. При полном отсутствии оледенений температуры воздуха и воды в океане были, вероятно, значительно выше современных: на экваторе на 3-5 °С, в средних широтах на 10 С, а в полярных - на 20-40 °С.
В то же время происходили существенные изменения рельефа Земли и положения континентов, которые привели к формированию в общих чертах существующих ныне континентов и океанов. В начале триаса сохранялись относительно высокие материки. Затем началась морская трансгрессия с образованием мелких краевых морей Тихого океана, прерывавшаяся в начале и в конце юрского периода Древне- и Позднекиммерийскими фазами горообразования. В меловом периоде новая морская трансгрессия привела к затоплению значительных территорий Европы и Северной Африки краевыми теплыми морями древнего океана Тетис.
По представлениям теории тектоники плит, общей тенденцией в мезозое был распад суперконтинента Пангеи и расхождения континентальных плит (рис. 60). В триасе этот процесс только начался. Лавразия сместилась несколько к северу. При этом происходило раскрытие океана Тетис, отделявшего Лавразию от Гондваны. Затем Лавразия раскололась на Северную Америку и Евразию, между которыми, начиная с раннеюрского времени, началось формирование северной части Атлантического океана. Вероятно, уже к концу юрского периода начался раскол Гондваны и, соответственно, возникновение Индийского океана и южной части Атлантики, отделивших Африку от Австралии и Антарктиды на востоке и юге и от Южной Америки на западе. Соединение северной и южной частей Атлантики произошло уже в позднемеловое время.
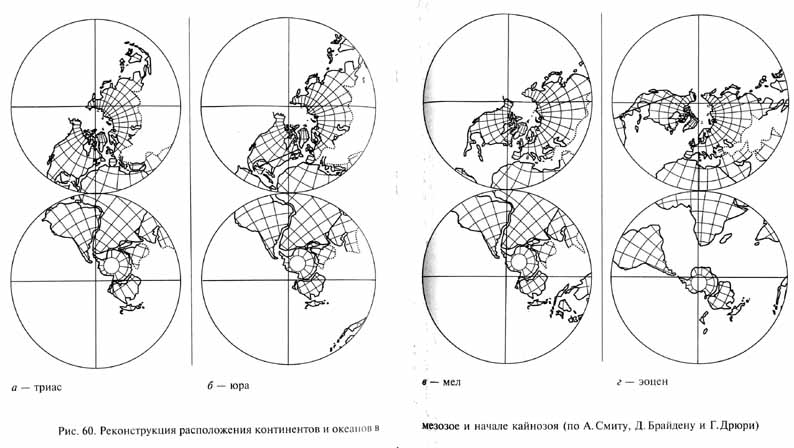
В меловом периоде расхождение континентов, которые в конце палеозоя входили в состав единой Пангеи, значительно прогрессировало. Продолжали расширяться Атлантический и Индийский океаны. При движении американских плит к западу на их западных окраинах возникли мощные зоны горной складчатости: в юрском периоде - Сьерра-Невада, в меловом - Анды и Скалистые горы (Позднекиммерийская и Ларамийская фазы горообразования). Индостан и Мадагаскар обособились от Африки, Индостанская плита начала смещение к северо-востоку. Сложные взаимные смещения Африки и Европы привели к постепенному закрытию океана Тетис. Из материков, входивших в состав Гондваны, в конце мезозоя остались соединенными лишь Антарктида и Австралия. Зато материки, составлявшие Лавразию и разделившиеся в начале мезозоя, к концу мела вновь соединились, правда противоположными концами: Азия и Северная Америка сошлись в области Чукотки и Аляски. Так возникла Берингия, сухопутный мост, связавший Америку и Европу на месте современного Берингова пролива.
Обновление флоры и фауны в триасе
Пермское оледенение и последовавшее после его завершения глобальное потепление климата привели к значительному изменению общего облика флоры. Вымерли древовидные плаунооб-разные, столь характерные для лесов второй половины палеозоя. Исчезли также семенные папоротники (птеридоспермы) и кордаиты. Их место занимают разнообразные представители других групп голосеменных растений: саговниковых (Cycadophyta), гинк-говых (Ginkgoales) и хвойных (Coniferales) (см. рис. 68, изображающий ландшафт юрского периода, где представлены указанные голосеменные, появившиеся уже в триасе). Особенно характерны для наземной флоры первой половины мезозоя были саговниковые: древовидные пальмообразные, с высокими колоннообразными стволами, увенчанными пучком перистых листьев (Williamsonia), или приземистые, с бочкообразным или клубневидным коротким и толстым стволом (Cycadeoidea). Саговниковые, хвойные и гинкговые формировали общий облик лесов в первой половине мезозойской эры.

Рубеж между пермским и триасовым периодами был отмечен также существенными изменениями в фауне наземных позвоночных. В пермское время преобладающими группами пресмыкающихся были зверообразные рептилии и котилозавры, тогда как группы с диапсидным черепом (лепидозавры и архозавры), объединяемые под названием "завропсиды", оставались сравнительно Немногочисленными. Это соотношение резко изменилось к началу мезозоя. Из числа котилозавров в триас перешли лишь проколофоны (вымирающие к концу этого периода), а из зверообразных - немногие группы дицинодонтов и высших териодонтов. Зато обилие и разнообразие диапсидных рептилий неуклонно возрас-Тает, и во второй половине триаса они становятся доминирую-Щими. При этом нужно отметить, что высшие териодонты обладали рядом прогрессивных признаков, отсутствовавших у завропсид (например, были способны к пережевыванию пищи, значительно повышающему степень ее усвоения организмом; вероятно, они имели теплоизолирующий волосяной покров и т.д.). Победа в борьбе за существование была достигнута завропсидами, так сказать, вопреки прогрессивным чертам организации териодонтов и, очевидно, основывалась на каких-то важных преимуществах первых.
П.Робинсон предложила гипотезу, связывающую изменения фауны рептилий в триасе с особенностями физиологии завропсид и зверообразных пресмыкающихся, о которых с известной долей вероятности можно судить по соответствующим характеристикам современных потомков тех и других. Современные завропсиды (в широком смысле этот термин используется для объединения всех современных групп этого класса и птиц) характеризуются наличием целого комплекса приспособлений к жизни в условиях жаркого засушливого климата. Завропсиды могут выдержать значительно более высокие температуры тела, чем потомки зверообразных рептилий - млекопитающие (многие ящерицы - до 44 °С, птицы - до 43 °С, млекопитающие - лишь до 39 °С; для Млекопитающих смертельно повышение температуры тела до 43 °С, Для птиц - до 46-47 °С). При этом рептилии используют энергию солнечного облучения для повышения температуры своего тела До оптимального уровня (гелиотермия); в связи с этим для них сохраняет важное значение "теменной глаз", регулирующий различные функции организма в зависимости от интенсивности падающего света.
Конечным продуктом белкового обмена у завропсид является мочевая кислота, тогда как у млекопитающих - мочевина. Мочевая кислота может образовывать перенасыщенные растворы, поэтому для ее выведения из организма требуется примерно в 10 раз меньше воды, чем для выведения мочевины. Очевидно, у завропсид выделительная система гораздо лучше экономит воду для организма, чем у млекопитающих.
Логично предположить, что организация завропсид складывалась под контролем отбора, приспосабливавшего этих рептилий к условиям засушливого и жаркого климата, тогда как зверообразные рептилии сформировались в местообитаниях с более влажным и прохладным климатом. Такие местообитания, вероятно, были широко распространены на континентах, входивших в состав Гондваны, в эпоху великого палеозойского оледенения. К концу пермского периода и в триасе во многих регионах климат становится все более засушливым (или, по крайней мере, характеризовался значительными сезонными засухами). П.Робинсон связывает это с изменениями морских течений и преобладающих направлений циркуляции атмосферных потоков в результате дрейфа континентов, входивших в состав Лавразии и Гондваны. Засушливость климата должна была дать важные преимущества в борьбе за существование тем группам наземных животных, которые обладали более высокой устойчивостью к действию высоких температур, были способны лучше экономить воду и с помощью специальных форм поведения могли использовать энергию солнечного облучения для достижения оптимальной температуры своего тела. Поскольку по всем этим показателям завропсиды превосходили зверообразных рептилий, последние были оттеснены в еще сохранившиеся более прохладные и влажные местообитания. В неблагоприятных для них условиях, при жесткой конкуренции со стороны завропсид, численность зверообразных рептилий значительно упала, и большинство их групп вымерло.
В немногих сохранившихся в триасе филогенетических стволах высших териодонтов продолжались изменения, наметившиеся еще среди пермских терапсид. Общее направление этих эволюционных изменений обозначают иногда термином "маммализация", т. е. развитие комплекса признаков, характерных для высших амниот -млекопитающих. Здесь мы вновь встречаемся с параллельной эволюцией: сходные черты строения независимо возникали в разных филетических линиях териодонтов. Среди этих признаков было постепенное увеличение размеров полушарий переднего мозга, приобретение мягких губ (что делало возможным сосание) и волосяного покрова. По мнению Л.П.Татаринова, зачаточные волоски у териодонтов имели осязательную функцию, располагаясь вблизи рта (как специализированные осязательные волосы - вибриссы у млекопитающих). Однако в среднетриасовых отложениях были обнаружены ископаемые следы цинодонта (Cynodontia - "собакозубые", одна из групп высших териодонтов) с отпечатками волос. Поэтому имеются основания считать, что у териодонтов волосяной покров был развит уже на всем теле, как у млекопитающих, и, вероятно, имел ту же основную функцию, как и у последних, т. е. служил для теплоизоляции (это понятно, если принять гипотезу о возникновении и развитии зверообразных рептилий в прохладном климате).
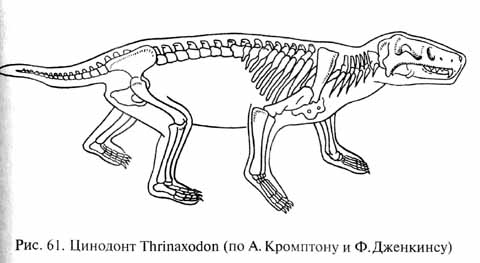
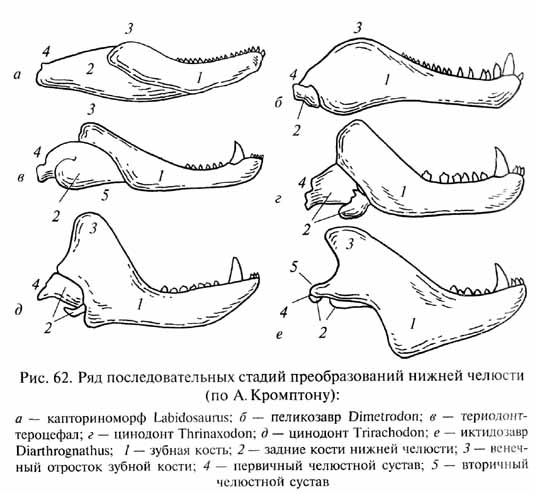
У цинодонтов (рис. 61) продолжалось совершенствование механизма пережевывания пищи с развитием окклюзии (смыкания коронок верхне- и нижнечелюстных зубов в щечной области) и продольных и поперечных движений нижней челюсти. Для обеспечения этих движений произошли сложные перестройки челюстных мышц, сыгравшие важную роль в дальнейшей эволюции челюстного аппарата. Механически наиболее благоприятное расположение челюстных мышц было достигнуто при разрастании зубной кости нижней челюсти назад и вверх с развитием высокого венечного отростка, к которому прикреплялась височная мышца (рис. 62). При этом задние кости нижней челюсти подверглись редукции. Гипертрофия задней части зубной кости Привела к возникновению ее контакта (при движениях нижней Челюсти) с чешуйчатой костью черепа. Между двумя этими костями возникла суставоподобная связь, которая оказалась механически более эффективной опорой и осью вращения для нижней челюсти, чем первичный челюстной сустав (между квадратной Костью верхней и сочленовной костью нижней челюсти). Преобразования нижней челюсти оказались преадаптивными для формирования новой опоры нижней челюсти - вторичного челюстного сустава.
Вторичный челюстной сустав, присущий среди всех позвоночных только млекопитающим, стал поэтому важнейшим диагностическим признаком последних. Редуцированные кости первичного челюстного сустава, освободившиеся от своей прежней функции, вошли в состав цепи косточек среднего уха в качестве двух наружных ее элементов - наковаленки и молоточка, что стало еще одной характерной особенностью млекопитающих, обеспечившей более тонкую слуховую чувствительность в области высоких звуковых частот.

Наиболее древние ископаемые остатки млекопитающих или зверей (Mammalia) известны из верхнетриасовых отложений. Это были небольшие хищные животные (рис. 63), питавшиеся различными мелкими позвоночными и беспозвоночными. Вероятно, они предпочитали прохладные и влажные местообитания и вели преимущественно ночной образ жизни. Скорее всего, у них уже существовало выкармливание детенышей молоком, поскольку оно присуще всем современным группам млекопитающих, а расхождение филогенетических стволов, ведущих, с одной стороны, к однопроходным (Monotremata), с другой - к сумчатым (Marsupialia) и плацентарным (Placentalia), произошло, возможно, уже в позднем триасе. Млечные железы возникли у древних млекопитающих из видоизмененных потовых желез. Возможно, первоначальной функцией млечных желез было не столько выкармливание, сколько "выпаивание" детенышей, т. е. снабжение их необходимой влагой и солями. Дефицит влаги был особенно опасен для новорожденных детенышей млекопитающих в условиях засушливого климата. Судя по способу развития однопроходных, наиболее примитивных среди современных млекопитающих, древнейшие звери еще были яйцекладущими животными, которые высиживали свою кладку или вынашивали яйца в специальной сумке на животе.
В конце триаса млекопитающие уже были довольно разнообразны и принадлежали к нескольким обособленным группам: три-конодонтам (Triconodontia), симметродонтам (Symmetrodontia) и харамиидам (Haramiyidae), различавшимся по строению зубов и, вероятно, по способам питания и пережевывания пищи.
Несмотря на ряд прогрессивных морфофизиологических особенностей (некоторое увеличение головного мозга, особенно его больших полушарий; забота о потомстве и выкармливание его молоком; пережевывание пищи; волосяной покров, расположение конечностей под туловищем, обеспечивающее более совершенную локомоцию), млекопитающие в мезозое не достигли больших успехов в конкуренции с рептилиями. Они оставались относительно малочисленными мелкими животными и, вероятно, вели ночной или сумеречный образ жизни, характерный для большинства зверей и ныне. Возможно, именно несовершенство систем терморегуляции и выделения, приспособленных к условиям относительно низких температур и высокой влажности, снижали для мезозойских млекопитающих шансы на успех в конкуренции с завропсидами. Хотя мезозойские млекопитающие уже обладали теплоизолирующим шерстным покровом и, вероятно, могли усилить теплоотдачу за счет потоотделения, у них не было еще совершенной интеграции разных терморегуляционных механизмов в единую систему. Об этом можно судить по состоянию современных однопроходных, у которых еще нет настоящей гомойотермии, и температура тела колеблется в пределах 26-34 °С.
В триасе начался подлинный расцвет рептилий. Особенно многочисленными становятся лепидозавры и архозавры. На ранних этапах их эволюционной истории эти два филогенетических ствола оставались еще очень близкими друг к другу, поэтому нередко трудно с достаточной уверенностью дифференцировать примитивных лепидозавров и архозавров. Таковы, например, ринхозавры (Rhynchosauria), которых одни авторы сближают с клювоголовы-ми среди лепидозавров, а другие относят к архозаврам. Ринхозавры получили широкое распространение в триасовое время. Это были крупные животные (1,5-5 м) с широким черепом, мошной жевательной мускулатурой и многорядными зубами на верхнечелюстных и зубных костях. Ринхозавры были растительноядными и, вероятно, конкурировали с дицинодонтами.
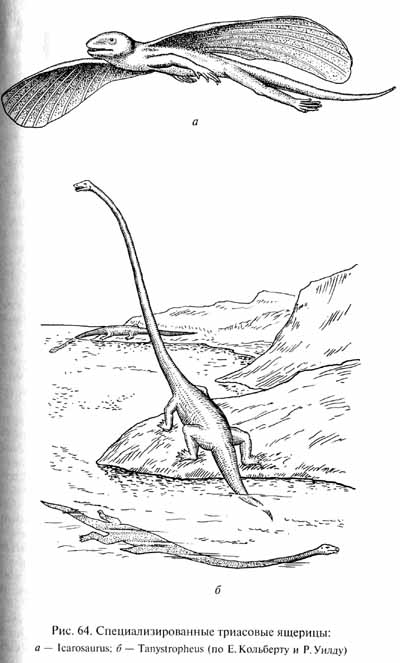
Настоящие клювоголовые (ринхоцефалы), родственные современной гаттерии, известны начиная с позднего триаса. В среднем триасе уже существовали ящерицы (группа эолацертилий), которые вскоре становятся очень разнообразными. Например, средне-триасовый танистрофей (Tanystropheus) обладал гипертрофированной шеей, длина которой превышала длину туловища (рис. 64, б)', общие размеры животного достигали 6 м. Возможно, эти рептилии, которых некоторые ученые также считают не ящерицами, а архозаврами, вели водный образ жизни, используя свою феноменальную шею при ловле рыб и головоногих моллюсков, как более поздние плезиозавры. Среди позднетриасовых эолацертилий имелись формы (Kuehneosaurus, Icarosaurus), приспособившиеся к планирующему полету: на удлиненных ребрах, выступавших по бокам туловища, была натянута летательная перепонка (рис. 64, а).
Возможно, вымирание последних котилозавров - проколофонов - в конце триаса было связано с их неудачей в конкуренции с экологически близкими, но более совершенными ящерицами.
Многочисленны и разнообразны в триасе были и представители архозавров. Корневой группой этого мощного филогенетического ствола являются псевдозухии, или текодонты (Pseudosi или Thecodontia), - преимущественно небольшие животные, примитивные представители которых внешне несколько напоминали ящериц. Но у псевдозухии череп сохранял типичное диапсидное строение, а зубы сидели в ячейках (текодонтный тип), а не прикреплялись к краю челюсти, как у ящериц. Для псевдозухии характерна тенденция к удлинению задних конечностей, связанная с переходом к быстрому бегу на двух ногах (рис. 65). Нужно отметить, что и ящерицы при максимальной скорости бегут лишь на двух задних конечностях, приподняв переднюю часть тела и длинный хвост над субстратом. Такой способ убыстрения бега (связанный с сохранением примитивной постановки конечностей, с горизонтальным положением плеча и бедра) был, видимо, характерен и для псевдозухии, но у последних получил более значительное развитие. Среди их потомков появились настоящие двуногие формы. Большинство псевдозухии были хищниками мелких и средних размеров (самые крупные достигали 3,5 м в длину). Некоторые из них стали быстро бегающими наземными животными, другие приспособились к лазанию по деревьям, были также полуводные формы - фитозавры (Phytosauria), имевшие внешнее сходство с крокодилами. В позднем триасе от псевдозухии возникли и настоящие крокодилы (Crocodylia), оказавшиеся лучше приспособленными к роли амфибиотических хищников и быстро вытеснившие фитозавров.

И фитозавры, и крокодилы конкурировали в водоемах с последними стегоцефалами - лабиринтодонтами, представленными в триасе очень крупными формами с огромным уплощенным черепом, достигавшим у Mastodonsauras 125 см длины. Однако и эти гиганты не выдержали конкуренции с водными рептилиями. Разнообразие и численность стегоцефалов в первой половине мезозоя неуклонно сокращались, и последние стегоцефалы (плагиозавры) вымерли в раннемеловое время.
В триасе появились также черепахи (Chelonia). Костно-роговой двуслойный панцирь черепах оказался замечательной защитной адаптацией, позволившей этим рептилиям сохранить многие примитивные черты организации, соответствующие уровню котилозавров. Происхождение черепах до сих пор остается дискуссионным. Древнейший известный их представитель, позднетриасовый Proganochelys, уже имел основные черты строения черепах. Разные исследователи видели предков черепах в различных группах котилозавров (от капториноморфов до парейазавров). М.Ф.Ивахненко, основываясь на некоторых особенностях строения черепа, высказал гипотезу о происхождении черепах от особого филогенетического ствола, названного Е.Олсоном парарептилиями и эволюционировавшего от уровня стегоцефалов независимо от других пресмыкающихся. Однако эта гипотеза требует допущения, Что все основные особенности амниот, рассмотренные нами выше (скелет, кожные покровы, дыхательная, кровеносная и выделительная системы, а также амниотический онтогенез), развивались независимо и параллельно у настоящих рептилий и парарептилий. Это представляется менее вероятным, чем параллельное (и неоднократное) формирование сходных особенностей черепа в разных линиях эволюции рептилий. Решение проблемы происхождения черепах, очевидно, ждет новых палеонтологических данных.
Жизнь в мезозойских морях
Теплые моря, заливы и лагуны в мезозое были богаты жизнью. После пермского вымирания разнообразие морских организмов в триасе вновь возрастает. По-видимому, в мезозое возник новый тип водорослей - диатомовые (Diatomeae). Во всяком случае, их ископаемые остатки достоверно известны, начиная лишь с юрских отложений.
В донной фауне опять обильны мшанки, многочисленны морские ежи и морские звезды. Постепенно возрастает численность и разнообразие брюхоногих моллюсков. Среди двустворчатых моллюсков выделяется мезозойская группа рудистов (Rudistae), прикреплявшихся к субстрату одной из створок своей асимметричной раковины, достигавшей у некоторых видов 1,5 м. В триасе начинается бурный расцвет аммонитов и белемнитов. Мезозойские головоногие были очень разнообразны и по форме раковины, и по размерам, варьировавшим у аммонитов от нескольких миллиметров до 3 м. Белемниты, подобно современным кальмарам, стремительно проносились в толще воды, охотясь за мелкими рыбами, а аммониты медленно плавали, регулируя глубину погружения с помощью раковины - гидростатического аппарата.
Постепенно обновилась ихтиофауна. В юрское время сформировались современные группы пластинчатожаберных хрящевых рыб (Elasmobranchia): акулы (Selachoidea) и скаты (Batoidea). Среди костных рыб появились новые группы лучеперых: костные ганои-ды (Holostei) и костистые рыбы (Teleostei). Первые пережили свой расцвет в юре, вторые возникли в юрское время, а начиная с мела стали доминирующей группой (к костистым принадлежит 95% всех современных видов рыб). Продолжали существовать в мезозойских морях кистеперые - целаканты.
Хрящевые рыбы (Chondrichthyes) полностью утратили костный скелет, характерный для всех древнейших палеозойских рыб. Это было, вероятно, связано с необходимостью освободиться от тяжелого наружного костного панциря при активизации образа жизни. Костные рыбы (Osteichthyes) также решали проблему облегчения костного панциря, но не путем его утраты вместе с внутренним костным скелетом, а посредством постепенных и более длительных преобразований, которые привели к формирований гибкого, легкого и прочного чешуйного покрова. Можно сказать, что в отношении преобразований наружного и внутреннего скелета хрящевые рыбы в сравнении с костными пошли по инадаптивному пути. В конкуренции с костными рыбами хрящевые рыбы компенсировали несовершенство своего скелета увеличением размеров тела и рядом прогрессивных изменений организации: совершенствование головного мозга, внутреннее осеменение, откладка крупных яиц, защищенных прочными оболочками, снабженных большим количеством желтка и обеспечивающих длительное развитие зародыша.
Всякая группа организмов, сумевшая в ходе эволюционных преобразований достичь нового уровня организации, под давлением естественного отбора начинает экологическую экспансию, осваивая все доступные местообитания, способы питания и т.п. При этом закономерно происходит адаптивная радиация - расхождение признаков у потомков общей предковой группы в процессе приспособительной эволюции, ведущее к возникновению разнообразных видов. Разнообразие развивающихся при этом приспособлений и возникающих жизненных форм зависит от совершенства и пластичности организации данной группы. Несомненно, одной из самых поразительных в этом отношении групп являются рептилии, которые в мезозое освоили практически все возможные типы местообитаний и дали фантастическое разнообразие жизненных форм, не превзойденное ни одним другим классом позвоночных животных.
Триас ознаменовался успешным освоением рептилиями не только пресных водоемов (танистрофеи, фитозавры и крокодилы), но и морей. Морские рептилии получили в мезозое широкое распространение и были чрезвычайно разнообразны: одни из них жили на мелководье в прибрежной зоне и, вероятно, сохраняли некоторую связь с сушей, другие же стали обитателями открытого океана и никогда не выходили на берег. Среди первых наиболее известны завроптеригии (Sauroptcrygia). Это были животные довольно странного облика (рис. 66): массивное широкое туловище, имевшее, однако, обтекаемую форму, впереди продолжал ось длинной и тонкой шеей (у некоторых форм она была длиннее туловища), а сзади - длинным сжатым с боков веслообразным хвостом, несущим сверху небольшой хвостовой плавник. Передние и задние конечности у более специализированных к жизни в море плезиозавров (Plesiosauria) имели строение широких и мощных ластов, которые были основными движителями в воде. Хвостовой Плавник был невелик и мог помогать при плавании с малой скоростью, а в основном он, вероятно, служил рулем. Ласты могли служить опорой при выходе плезиозавров на прибрежные отмели Для откладки яиц. как это делают современные морские черепахи. Однако даже такое кратковременное возвращение на сушу для Некоторых плезиозавров было очень затруднено их огромными размерами: ряд юрских и меловых видов достигал 12-16 м (плиозавры, элазмозавры).
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 1317; Нарушение авторских прав?; Мы поможем в написании вашей работы!