КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Часть III. Историческое развитие (филогенез) организмов 1 страница
|
|
|
|
Анализ развития приспособлений организмов подводит нас к новому кругу проблем, относящихся уже к уровню макроэволюции. Собственно говоря, такие аспекты адаптациогенеза, как морфофункциональные преадаптации, инадаптивная эволюция, гиперадаптивность, мы по необходимости уже рассматривали в широкой исторической перспективе, т.е. именно в масштабе макроэволюции. С этими вопросами тесно связаны специфические проблемы макроэволюции: механизмы типогенеза (формирование основного плана строения, или типа организации высших таксонов) и типостаза (последующая длительная устойчивость этого общего плана организации), направленность эволюционного процесса и др. Чтобы разобраться в этих сложных проблемах, целесообразно предварительно рассмотреть реалии макрофилогенеза: его исторические масштабы, процессы формирования и историческую судьбу основных групп организмов, причины их расцвета и вымирания, соотношения этих процессов с изменениями среды обитания в геологической истории. В сущности, эволюция организмов представляет собой форму существования живой материи в изменяющейся с течением времени внешней среде, и все современные проявления жизни на любом уровне организации живой материи могут быть поняты лишь с учетом их эволюционной истории. Эти проблемы и рассматриваются в данной части книги.
Однако прежде чем мы приступим к рассмотрению последовательных этапов эволюции органического мира, нам необходимо отчетливо представлять временные масштабы этих процессов, т. е. масштабы геологического времени. Длительное и кропотливое изучение абсолютного и относительного возраста горных пород в разных регионах земного шара, потребовавшее работы нескольких Поколений геологов и палеонтологов, позволило наметить основные вехи геологической истории. История Земли разделена на крупные временные промежутки, называемые геологическими эрами; эры (за исключением наиболее древних) разделены на геологические периоды, а те, в свою очередь, - на эпохи (табл. 2). Границы Между этими подразделениями соответствуют разного рода изменениям геологического и биологического (палеонтологического) Характера: усиление вулканизма и горообразовательные процессы; поднятия или опускания значительных участков континентальной коры, приводившие к соответствующим вторжениям или отступаниям моря (морские трансгрессии и регрессии); существенные изменения фауны и флоры и т.п. Поскольку подобные события происходили в истории Земли нерегулярно, продолжительность различных эпох, периодов и эр различна.
Отложения древнейших археозойской и протерозойской эр содержат чрезвычайно мало ископаемых остатков организмов; по этому признаку археозой и протерозой нередко объединяют под названием "криптозой" (этап скрытой жизни), противопоставляя трем последующим эрам - палеозою, мезозою и кайнозою, объединяемым как "фанерозой" (этап явной, наблюдаемой жизни).
Таблица 2. Основные подразделения геологической истории Земли, их абсолютный возраст и продолжительность (в млн. лет):
В последние десятилетия в позднепротерозойских (вендских) отложениях были сделаны довольно многочисленные находки ископаемых остатков разнообразных многоклеточных животных ("Эдиакарская фауна", см. с. 157). Поэтому некоторые ученые считают целесообразным присоединить венд к фанерозою.
Возраст Земли определяется различными учеными по-разному; можно указать приближенную цифру 5 млрд лет. Древнейшие известные на Земле горные породы - граниты Кольского полуострова - имеют возраст (установленный радиоактивным методом) не менее 4 млрд лет.
ГЛАВА 1. РАЗВИТИЕ ЖИЗНИ В КРИПТОЗОЕ И ЗАГАДКА НИЖНЕГО КЕМБРИЯ
Относящиеся к криптозою археозойская и протерозойская эры вместе продолжались более 3,4 млрд лет, тогда как три эры фанерозоя - 570 млн лет, т.е. криптозой составляет свыше 85 % всей геологической истории. Однако в отложениях криптозоя сохранилось чрезвычайно мало ископаемых остатков организмов (всего около 3000 местонахождений на 3,4 млрд лет), поэтому наши представления о первых этапах развития жизни в течение этих огромных промежутков времени в значительной степени гипотетичны.
Развитие жизни в первой половине криптозоя
Предполагаемые древнейшие остатки организмов были описаны из археозойских пород с возрастом 3,1-3,8 млрд лет из различных районов земного шара (Гренландия, Южная Африка, Западная Австралия). Из этих находок лучше известны ископаемые остатки, обнаруженные в кремнистых сланцах системы Свазиленд (Восточный Трансвааль, Южная Африка). В этих осадочных породах встречаются различные микроскопические структуры, которые рассматриваются как ископаемые остатки одноклеточных сине-зеленых водорослей (Archaeospheroides barbertonensis) и бактерий (Eobacterium isolatum). Первые имеют сфероидную форму диаметром 17-20 мкм, вторые - палочковидную длиной 0,5-0,7 мкм и толщиной 0,2-0,3 мкм. Отмечены также нитчатые формы, напоминающие нитчатые синезеленые водоросли. Геохимическими методами было показано наличие в осадочных породах, вмещающих эти ископаемые остатки, ряда органических веществ, которые, судя по содержанию в них разных изотопов углерода, могут иметь биогенное происхождение, некоторые из них могут быть производными хлорофилла. Это указывает на возможность существования фотосинтезирующих организмов уже 3,2 млрд лет назад.
Необходимо оговориться, что принадлежность археозойских ископаемых остатков к древнейшим организмам не может пока считаться вполне доказанной. Некоторые ученые полагают, что это - псевдоископаемые, образовавшиеся абиогенным путем'. Структурно археозойские ископаемые остатки очень просты, поэтому пока нет оснований утверждать, что они не могли возник-луть абиогенно.
Однако в осадочных породах Западной Австралии, имеющих возраст 3,4-3,5 млрд лет, вместе с остатками микроорганизмов, подобных таковым из южноафриканских местонахождений, были обнаружены древнейшие строматолиты - следы жизнедеятельности водорослей (вероятно, синезеленых). Строматолиты найдены также в несколько более молодых отложениях Южной Африки в Зимбабве возрастом 2,9-3,2 млрд лет. Все эти находки убедительно свидетельствуют, что около 3,3 млрд лет назад на Земле уже существовали водоросли. Очевидно, появление жизни на Земле должно было произойти раньше - может быть, 4-4,25 млрд лет назад.
Протоорганизмы были, вероятно, гетеротрофными формами, которые питались готовыми высокомолекулярными органическими соединениями, содержавшимися в первобытном океане ("первичном бульоне") и имевшими абиогенное происхождение. В археозое жизнь существовала в условиях бескислородной восстановительной атмосферы, вероятно, в водоемах на глубине порядка 10-50 м. Десятиметровый слой воды защищал протоорга-низмы от губительного действия жесткого ультрафиолетового излучения Солнца.
В промежутке времени между 3,3 и 3,5 млрд лет какие-то формы древнейших организмов - прокариот, еще не имевших ядра, приобрели способность к фотосинтезу. В связи с этим произошла дивергенция прокариот на два главных ствола их эволюции:
1) бактерии (Bacteriae), в большинстве сохранившие в той или иной форме гетеротрофный способ питания и совершенствовавшие его применительно к новым условиям существования, и
2) синезеленые водоросли (Cyanophyta), развившие автотрофное питание посредством фотосинтеза. Бактерии от использования органических веществ абиогенного происхождения по мере уменьшения их содержания в окружающей среде и распространения фотосинтезирующих организмов все в большей степени переходили к питанию органическими веществами, синтезированными водорослями.
В течение последующих 1,5-2 млрд лет - на протяжении большей части криптозоя - происходила эволюция прокариот, в ходе которой возникли новые варианты их строения, метаболизма, питания, но основные их черты, характеризующие прокариотический уровень организации, оставались неизменными. Вероятно, около 3,3 млрд лет назад появились нитчатые формы синезс-леных водорослей. Нитчатый тип строения представляет собой цепочку клеток, возникшую путем их последовательного деления в одном направлении. Клетки в цепочке связаны друг с другом лишь механически -- разрыв цепочки никак не сказывается на жизнедеятельности и жизнеспособности составляющих ее клеток. Поэтому нитчатые формы нельзя приравнивать к многоклеточному состоянию, характеризующемуся дифференциацией и интеграцией целостного организма, различные части которого выполняют разные функции и подчинены целому.
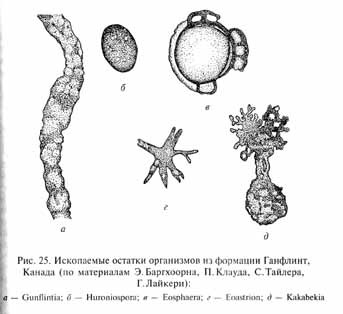
В отложениях первой половины протерозоя (возрастом 2,5- 1,7 млрд лет) ископаемых остатков организмов несколько больше, а главное - их природа не вызывает таких сомнений, как для более древних микроископаемых. Наиболее известны ископаемые остатки флоры, обнаруженные в Канаде в кремнистых сланцах формации Ганфлинт на северном берегу озера Верхнего. Возраст этих отложений составляет около 1,9 млрд лет. В строматолитовых кремнистых породах здесь встречаются ископаемые остатки довольно разнообразных микроорганизмов (рис. 25). Обычны нитчатые формы длиной до нескольких сот микрометров (микронов) и толщиной 0,6-16 мкм различного строения: простые и ветвистые, нечленистые и разделенные по длине на сегменты (клетки?), причем последние могут быть разной формы - округлые и удлиненные. Строение некоторых нитевидных организмов (Gunflintia), по мнению Е. Баргхоорна, напоминает таковое современных синезеленых водорослей, другие же сходны с железобактериями. Имеются и одноклеточные микроорганизмы диаметром 1-16 мкм, также различного строения: от простых округлых, снабженных толстой оболочкой Huroniospora, до сложных причудливой формы Eoastrion и Kakabekia. В этих породах обнаружены и молекулярные ископаемые - органические вещества, подобные найденным в более древней формации Свазиленд, но более разнообразные.
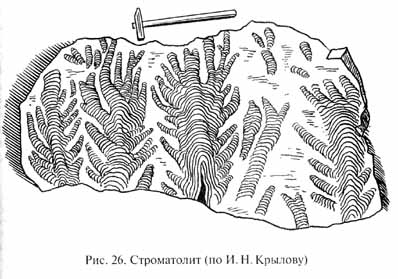
В осадочных породах, образовавшихся в промежутке времени между 2 и 1 млрд лет назад, часто встречаются строматолиты (рис. 26), что говорит о широком распространении и активной фотосинтезирующей деятельности синезеленых водорослей в этот период.
Жизнь в среднем протерозое и происхождение эукариот
Следующий важнейший рубеж в эволюции жизни документируется рядом находок ископаемых остатков в отложениях, имеющих возраст 0,9-1,3 млрд лет. Среди них наиболее известна ископаемая флора, обнаруженная в углистых кремнистых сланцах формации Биттер-Спрингс (возраст 0,9 млрд лет) в Центральной Австралии. Помимо одноклеточных и нитчатых форм синезеленых водорослей (по мнению Д. Шопфа, они уже относятся к современным семействам этих растений) здесь найдены прекрасной сохранности остатки одноклеточных организмов размером 8-12 мкм, в которых удалось различить внутриклеточную структуру, похожую на ядро (рис. 27). Обнаружены также стадии деления одного из видов этих одноклеточных организмов (Glenobotrydion aenigmati-cus), напоминающие стадии митоза - способа деления эукарио-тических (т.е. имеющих ядро) клеток. Сходные ископаемые остатки содержащих ядро клеток и стадий митоза были найдены также в Калифорнии в отложениях, имеющих возраст 1,3 млрд лет, а также на реках Мироедиха (Туруханский район) и Мая (Юго-Восточная Якутия) в отложениях возрастом 0,9-1 млрд лет.
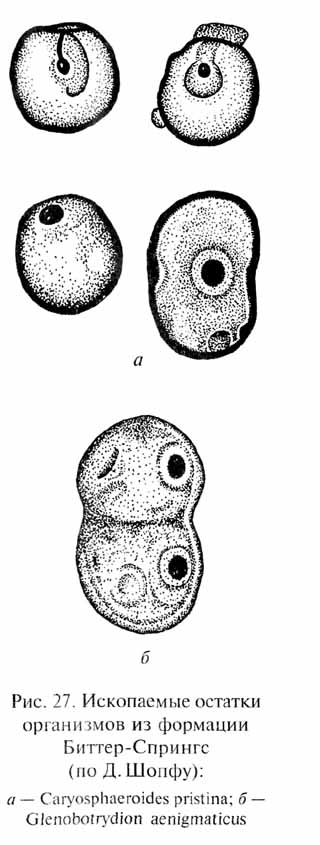
Если интерпретация всех этих ископаемых остатков правильна1, это означает, что около 1,6-1,3 млрд лет назад эволюция организмов прошла важнейший рубеж - был достигнут уровень организации эукариот.
Ранний протерозой с полным основанием называют "веком синезеленых водорослей", получивших самое широкое распространение; это было время их расцвета. Благодаря их фотосинтетической деятельности постепенно повышалось содержание кислорода в атмосфере. Первоначально свободный кислород быстро использовался как окислитель в различных химических реакциях, в частности в процессах отложения осадочных железных руд при участии ферробактерий (основная масса железорудных толщ на всей Земле сформировалась в промежутке 2,2-1,9 млрд лет назад). После завершения этих процессов, радикально изменивших геохимический облик поверхностных слоев земной коры, кислород стал накапливаться в атмосфере во все более заметных количествах. Большие масштабы в это время приобрела и строматолитообразующая деятельность нитчатых синезеленых водорослей, которая привела к образованию огромных толщ карбонатных пород.
Возникновение эукариот - это следующий важнейший рубеж в до-кембрийской эволюции организмов после возникновения фотосинтеза. Эукариоты обладают значительно более сложной и совершенной организацией клетки, чем прокариоты. Протоплазма эукариотической клетки сложно дифференцирована: в ней обособлены ядро и другие различные органоиды (митохондрии, пластиды, комплекс Гольджи, центриоли и др.), отделенные от цитоплазмы полупроницаемыми мембранами и выполняющие различные функции, снаружи клетка окружена клеточной мембраной. Наружная и внутренние клеточные мембраны образуют единый мембранный комплекс. Мембраны имеют сложное молекулярное строение (липидная "пленка", пронизанная белковыми "каналами"), обусловливающее избирательное проникновение веществ. У прокариот вместо клеточной мембраны клетка обволакивается единственной гигантской молекулой мукопептида (вещество, состоящее из аминокислот, углеводов и липидов). В ядре - интегрирующем центре клетки эукариот - локализован хромосомный аппарат, в котором сосредоточена основная часть
кода наследственной информации. У прокариот нет такого интегрирующего центра и подобной упорядоченности наследственной информации; элементы аппарата наследственности отчасти включены в единственную так называемую "хромосому" бактериальной клетки, отчасти находятся в цитоплазме. Упорядоченность аппарата наследственности и сложность организации клетки у эукариот требуют такой же упорядоченности в передаче наследственной информации при клеточном делении. В связи с этим у эукариот развился весьма совершенный механизм клеточного деления - митоз, обеспечивающий точное и эквивалентное распределение наследственной информации в обе дочерние клетки. У прокариот митоза не наблюдается.
Одним из важнейших эволюционных достижений эукариот является возникновение механизма цитоплазматической подвижности, связанное с формированием аппарата микротрубочек (и его производных), в котором локализованы сократительные белки: миозин, актин, тубулин (у прокариот актин и тубулин встречаются редко, а миозин неизвестен). Источником энергии для функционирования сократительных белков является аденозинтрифосфат (АТФ) - основной аккумулятор и переносчик химической энергии в клетке. Возникновение митоза стало возможным именно благодаря развитию этого аппарата, с ним же связано и совершенствование механизмов активной подвижности организма во внешней среде. (Среди прокариот подвижностью обладают некоторые жгутиконосныс бактерии. Однако жгутики бактерий не способны изгибаться. Они образованы белком флагеллином, не обладающим АТФ-азной активностью. Источником энергии для движения бактерий является градиент ионов водорода на бактериальной мембране).
Эукариотный уровень организации обеспечивает более эффективное выполнение всех клеточных функций и открывает перспективы для дальнейшей прогрессивной эволюции.
Имеются две основные точки зрения на проблему происхождения эукариотической клетки и соответственно эукариот как одной из высших таксономических групп. Первая из них - так называемая аутогенная концепция - предполагает постепенную дифференциацию прокариотической клетки, в ходе которой развился мембранный комплекс. Сначала могла возникнуть наружная клеточная мембрана, затем ее локальные впячивания внутрь клетки образовали внутриклеточные мембраны. На основе дифференциации мембранного комплекса структурно оформились клеточные органоиды. Согласно аутогенной концепции эукариоты возникли от одной из групп прокариот (от какой именно - указать сейчас невозможно).
Вторая точка зрения (симбиогенная концепция) получила широкую известность после работ Л.Маргулис (Саган), в 1967-1971 гг. обосновавшей гипотезу о возникновении эукариотической клетки из симбиоза разных прокариот: крупной клетки - хозяина - и более мелких организмов, поселившихся в цитоплазме первого. Последние дали начало различным органоидам. Аргументы в пользу этой точки зрения сводятся к наличию в некоторых органоидах эукариотической клетки (митохондрии, пластиды) собственной ДНК, некоторому сходству нуклеотидных последовательностей этих органоидов с таковыми ряда прокариот, а также су-шествованию сложных симбиотических комплексов среди современных организмов (известно, например, около тысячи видов-эндосимбионтов, обитающих в организме одноклеточных простейших животных). Согласно симбиогенной концепции предками эукариот должны быть сразу несколько групп прокариот.
Симбиогенная концепция получила широкое распространение в течение двух последних десятилетий. Однако все же аутогенная (классическая) точка зрения представляется в настоящий момент лучше обоснованной и согласующейся с общими тенденциями прогрессивной эволюции организмов (см. ч. IV). ДНК органоидов могла возникнуть путем сегрегации (обособления) фрагментов первоначально единого генома.
Для эукариот характерен половой процесс, сущностью которого является обмен наследственной информацией между организмами, принадлежащими к одному биологическому виду. Как мы видели выше, половой процесс объединяет наследственную информацию, присущую разным индивидам, в единый видовой генофонд и, с другой стороны, обеспечивает перекомбинацию наследственной информации в каждом следующем поколении. Половой процесс известен и у некоторых прокариот (бактерии), но у них он неупорядочен и не обеспечивает эквивалентного обмена разных особей наследственной информацией.
Вероятно, практически одновременно с развитием у эукариот полового процесса у них должен был сформироваться и мейоз, необходимый для редукции числа геномов, удваивающегося при половом процессе. Мейоз также способствует возрастанию комбинативной изменчивости вида. Результатом ее резкого усиления у эукариот было значительное убыстрение их эволюции. Вместе с тем организация клеточного ядра и аппарата клеточного деления 1у эукариот обеспечивает гораздо более совершенную защиту генома от горизонтального переноса генов, чем это характерно для прокариот. Горизонтальный перенос генов для эукариот теряет существенное значение как фактор эволюции.
Уже около 0,9 млрд лет назад произошло разделение эволюционного ствола древнейших эукариот на ряд ветвей, давших начало разным типам водорослей: зеленым (Chlorophyta), бурым (Phaeophyta), красным (Rhodophyta) и др., а также грибам (Fungi). Д.Шопф полагает, что описанные выше ископаемые остатки различных эукариот из местонахождений возраста 0,9-1,3 млрд лет принадлежат ко всем этим группам низших растений.
Вероятно, в это время существовали уже и одноклеточные животные -- простейшие (Protozoa), либо возникшие от общего Предкового ствола всех эукариот, либо обособившиеся от ранних Представителей одной из групп растений. От какой именно группы - с уверенностью сказать сейчас невозможно: разные исследователи связывали происхождение животных с каждой из названных групп растений. По общей совокупности данных, к предкам Protozoa наиболее близкими кажутся некоторые одноклеточные зеленые водоросли. Не случайно таких жгутиконосных одноклеточных, как эвгленовые, вольвоксовые и др., способных и к фотосинтезу, и к гетеротрофному питанию, ботаники традиционно рассматривают в составе типа зеленых водорослей, а зоологи - в составе типа простейших животных.
Обращает на себя внимание отсутствие ископаемых остатков животных в отложениях большей части криптозоя. Остатки некоторых одноклеточных простейших (радиолярии и, возможно, фо-раминиферы) и ряда групп многоклеточных животных появляются лишь в вендских отложениях. Это может быть связано и с более поздним возникновением животных, и с худшими возможностями фоссилизации (т.е. перехода в ископаемое состояние) их клеток, лишенных в отличие от растительных прочной целлюлозной оболочки. Скелета же протерозойские животные не имели.
Происхождение многоклеточных организмов
Вероятно, 700-900 млн лет назад на Земле появились первые многоклеточные животные и растения. У растений возникновение многоклеточного уровня организации, вероятно, произошло на основе дифференциации лентообразных колоний, образовавшихся путем бокового срастания прикрепленных нитчатых форм или благодаря делению клеток последних в двух взаимно перпендикулярных направлениях (в одной плоскости). У колоний, прикрепленных одним концом к субстрату, различные участки находились в разных условиях по отношению к падающему свету, субстрату и водной среде. В связи с этим естественный отбор благоприятствовал возникновению некоторой дифференциации частей колонии. Первым шагом было возникновение полярности колонии; на одном ее конце выделялись клетки, служившие для прикрепления к субстрату (для них характерно ослабление фотосинтеза, потеря способности к делению), на другом же конце - верхушечные клетки, интенсивно делившиеся и образовавшие своего рода "точку роста" колонии. Естественный отбор благоприятствовал приобретению клетками колонии способности делиться в разных направлениях; это приводило к ветвлению, что увеличивало поверхность колонии. Деление клеток вдоль трех взаимно перпендикулярных осей или переплетение отдельных нитей вело к возникновению многослойного "объемного" тела. В процессе его дальнейшей дифференциации сформировались многоклеточные органы, выполнявшие разные функции (фиксация на субстрате, фотосинтез, размножение). Одновременно между разными клетками растения складывалась определенная взаимозависимость, что, собственно говоря, и знаменует достижение многоклеточного уровня организации.
У животных активный образ жизни требовал более совершенной и сложной дифференциации организма, чем у растений. Сложность организации многоклеточных животных (Metazoa) и разнообразие ее конкретных форм стимулировали разработку различных гипотез о происхождении Metazoa.
Первая из них берет начало в работах Э.Геккеля, который в разработке своей теории гастреи основывался на сформулированном им биогенетическом законе, согласно которому онтогенез данного вида организмов представляет собой сжатое и сокращенное повторение (рекапитуляцию) хода филогенеза его предков (подробнее см. в ч. IV). В соответствии с этим Э.Геккель полагал, что филогенез древнейших Metazoa в определенной степени повторяется в онтогенезе современных низших многоклеточных животных (рис. 28). Согласно Геккелю, предками Metazoa были колониальные простейшие, обладавшие сферическими колониями с однослойной стенкой, подобными бластуле - одной из ранних стадий эмбрионального развития современных многоклеточных животных. Геккель назвал эту гипотетическую предковую форму "бластеей". При направленном плавании сферическая колония - бластея - ориентировалась одним полюсом вперед, как это наблюдается и у современных колониальных простейших, например у Volvox. Согласно Геккелю, на переднем полюсе колонии возникло впячивание ее стенки внутрь, подобно тому, как это происходит при инвагинационной гаструляции в онтогенезе некоторых современных Metazoa. В результате образовался многоклеточный организм - "гастрея", стенка тела которого состоит из двух слоев, экто- и энтодермы. Энтодерма окружает внутреннюю полость - первичный кишечник, открытый наружу единственным отверстием - первичным ртом. Организация гастреи соответствует принципиальному плану строения кишечнополостных (тип Coelenterata), которых Геккель и рассматривал как наиболее примитивных многоклеточных животных.
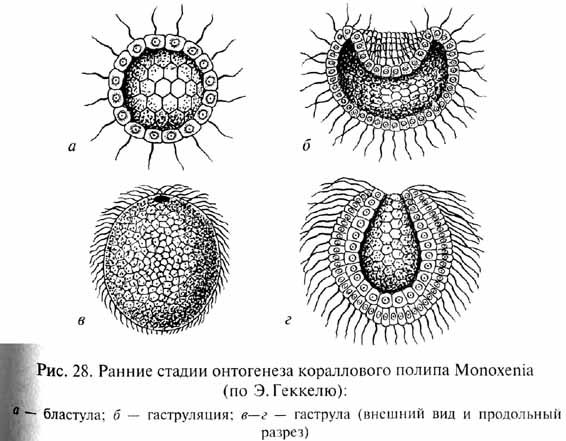
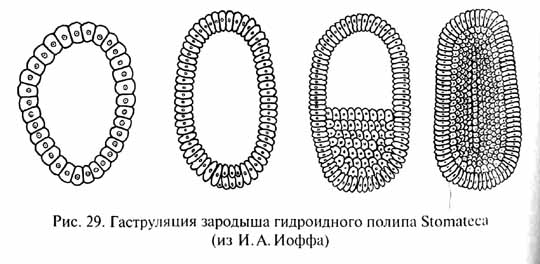
И.И.Мечников обратил внимание на то, что у примитивных кишечнополостных гаструляция происходит не путем инвагинации (впячивания одного полюса однослойного зародыша - бластулы), что характерно для более высокоорганизованных групп, а посредством миграции некоторых клеток из однослойной стенки тела внутрь (рис. 29). Там они образуют рыхлое скопление, позднее организующееся в виде стенок гастральной полости, которая прорывается наружу ротовым отверстием. Такой способ гаструляции гораздо проще, чем инвагинация, так как не требует сложного направленного и координированного смещения целого пласта клеток, и, вероятно, примитивнее инвагинации. В связи с этим Мечников модифицировал гипотезу Геккеля следующим образом. В сфероидной колонии простейших - жгутиконосцев клетки ее однослойной стенки, захватывавшие (фагоцитировавшие) пищу, мигрировали для ее переваривания внутрь, в полость колонии (подобно миграции клеток будущей энтодермы в Процессе гаструляции кишечнополостных). Эти клетки образовали рыхлое внутреннее скопление -- фагоцитобласт, функцией которого стало обеспечение всего организма пищей, включая ее переваривание и распределение, тогда как поверхностный слой клеток - кинобласт - осуществлял функции защиты и движения организма. Для захвата новых пищевых частиц клеткам фагоци-тобласта, по мысли Мечникова, не было необходимости возвращаться в поверхностный слой: располагаясь непосредственно под кинобластом, клетки фагоцитобласта захватывали пищевые частицы псевдоподиями, выдвигаемыми наружу в промежутках между клетками фагоцитобласта. Эта гипотетическая стадия эволюции Metazoa была названа Мечниковым фагоцителлой (или паренхимеллой); ее строение соответствует таковому паренхимулы, личинки некоторых кишечнополостных и губок. В дальнейшем как приспособление к повышению активности питания у потомков фагоцителлы произошла эпителизация фагоцитобласта с образованием первичного кишечника и возникновением ротового отверстия в том месте, где происходила преимущественная миграция клеток внутрь. По мнению некоторых ученых, это место, вероятно, соответствовало заднему по направлению движения полюсу тела, где при плавании возникают завихрения водяного потока, и поэтому условия наиболее благоприятны для захвата пищевых частиц. Гипотеза Мечникова, как и гипотеза Геккеля, рассматривает в качестве наиболее примитивных многоклеточных животных кишечнополостных и губок.
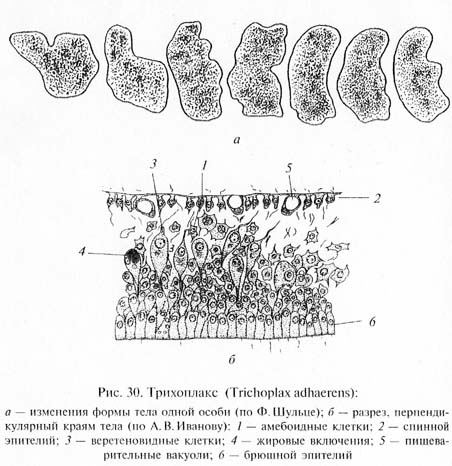
Важные сведения для понимания ранних этапов эволюции Metazoa были получены при изучении крайне примитивного многоклеточного животного трихоплакса (Trichoplax adhaerens), обнаруженного в Красном море Ф. Шульце еще в 1883 г., но детально исследованного лишь в 1970-е гг. нашего века К. Греллом и А. В. Ивановым. Трихоплакс (рис. 30) имеет уплощенное тело, лишенное полярности. Поверхность тела, обращенная вверх, покрыта плоским, а нижняя - цилиндрическим мерцательным эпителием. Внутри, между эпителиальными слоями, соответствующими кинобласту, находится полость с жидким содержимым, в Котором располагаются веретеновидные и звездчатые клетки. Эти Последние можно рассматривать как фагоцитобласт. Размножайся трихоплакс бесполым способом - делением и почкованием. А.В. Иванов указал, что трихоплакс представляет собой как бы Живую модель фагоцителлы, и предложил выделить эту форму в °собый тип животных Phagocytellozoa. По-видимому, трихоплакс Подкрепляет позиции гипотезы фагоцителлы И.И.Мечникова. Однако по современным представлениям непосредственными потомками фагоцителлозой среди многоклеточных были не кишечнополостные, а примитивные червеобразные животные, близкие по уровню организации к плоским ресничным червям - турбелляриям.
Первые ископаемые следы жизнедеятельности червеобразных многоклеточных животных известны из позднерифейских отложений. В вендское время (650-570 млн лет назад) существовали уже разнообразные животные, вероятно принадлежавшие к различным типам. Немногочисленные отпечатки мягкотелых вендских животных известны из разных районов всех континентах земного шара, кроме еще малоисследованной Антарктиды. Ряд интересных находок был сделан в позднепротерозойских отложениях на территории России -- на Кольском полуострове, Архангельской области, на реке Мая и на Оленекском поднятии в Якутии и т.д.
Наиболее известна богатая позднепротерозойская фауна, обнаруженная в Центральной Австралии в районе Эдиакары к северу от г.Аделаида. Исследовавший эту фауну М.Глесснер считает, что она включает несколько десятков видов очень разнообразных многоклеточных животных, относящихся к разным типам (рис. 31). Большинство форм принадлежат, вероятно, к кишечнополостным. Это медузоподобные организмы, вероятно "парившие" в толще воды (Ediacara flindersi, Beltanella gilesi, Medusinites asteroides и др.), и прикрепленные к морскому дну полипоидные формы, одиночные или колониальные, напоминающие современных кораллов альционарий, или морские перья (Rangea longa, Arborea arboкa, Pteridinium simplex и др.). Замечательно, что все они, как и Другие животные эдиакарской фауны, лишены твердого скелета.
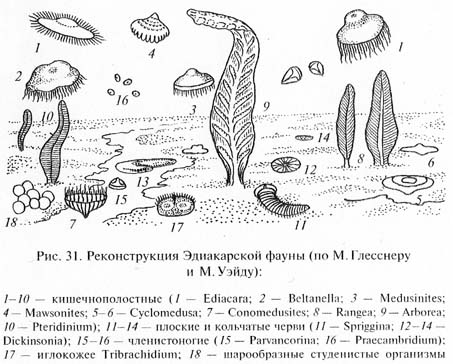
Кроме кишечнополостных в составе эдиакарской фауны найдены остатки червеобразных животных, причисляемых к плоским и кольчатым червям (Spriggina flounderi и разные виды Dickinsonia). Некоторые виды организмов интерпретируют как возможных пред-Ков членистоногих (Praecambridium sigillum, напоминающий по Характеру сегментации тела трилобитов и хелицеровых) и иглокожих (Tribrachidium heraldicum с телом дисковидной формы, на плоской поверхности которого выступают три валика, и Arkaria adami с пятилучевой звездообразной впадиной на ротовой стороне тела и с подобием амбулакральных желобков). Наконец, имеется целый ряд ископаемых организмов неизвестной таксономической принадлежности.
Многие вендские организмы были обнаружены также в вендских отложениях разных районов России: медузоподобные эдиакария и медузинитес - на полуострове Рыбачьем, птеридиниум - на севере Якутии, напоминающая сприггину вендия - в районе Яренска Архангельской области и т.д. Местонахождения вендской фауны, по богатству не уступающие эдиакарскому, были найдены на реке Сюзьма на Онежском полуострове и на Зимнем берегу Белого моря. Здесь были обнаружены ископаемые остатки свыше 30 видов бесскелетных многоклеточных животных, размеры которых варьировали от 3 мм до 30 см. Среди них вероятные представители кишечнополостных, плоских и кольчатых червей, членистоногих, иглокожих, а также ряд форм, принадлежащих к каким-то неизвестным группам. Вообще родство вендских организмов с современными группами, большинство из которых достоверно известно начиная с кембрия, остается проблематичным - различия очень велики, и некоторые исследователи считают, что известные ныне вендские организмы не связаны прямым родством с более поздними кембрийскими, а представляют слепые эволюционные ветви.
М.А.Федонкин, исследовавший Беломорскую фауну вендских животных, полагает, что некоторые из этих организмов обладают признаками нескольких разных типов животных и могут представлять исходные формы, занимающие промежуточное положение. Федонкин обратил также внимание на сходство ряда вендских организмов с личиночными стадиями некоторых современных животных, хотя вендские организмы имеют значительно более крупные размеры, чем соответствующие личинки. При всем разнообразии планов строения тела у организмов вендской фауны ("вендобионтов") их объединяют некоторые общие черты организации: отсутствие скелета, конечностей, вероятно, также дыхательных и пищеварительных органов. Многие вендобионты вели неподвижный прикрепленный образ жизни. Некоторые исследователи полагают, что вендские организмы питались осмотически через поверхность тела или же с помощью живших в их теле фото- или хемосинтезируюших симбионтов - одноклеточных водорослей и бактерий.
Хотя среди вендских животных преобладают мягкотелые бесскелетные формы, вероятно, в те времена существовали уже и немногие виды, обладавшие раковиной. Такова, например Cloudina, имевшая простую трубчатую раковину, состоявшую из органического вещества и кальцита. Клаудина была обнаружена в карбонатных породах, которые переслаиваются с отложениями, содержащими остатки эдиакарской фауны мягкотелых животных.
Все эти данные указывают на широкое распространение фаун мягкотелых животных в вендское время. Накопление материалов по вендским ископаемым организмам позволило некоторым исследователям поставить вопрос о расширении рамок фанерозоя, с включением в его состав "эдиакария" - периода, охватывающего промежуток времени от 670 до 550 млн лет назад (в приведенной на с. 143 схеме геохронологии этот период соответствует венду в составе протерозоя).
Поскольку вендская фауна столь разнообразна и включает довольно высокоорганизованных животных, очевидно, что до ее возникновения эволюция Metazoa продолжалась уже очень долго. Вероятно, многоклеточные животные появились значительно раньше - где-то в промежутке 700-900 млн лет назад. (Недавно появились первые данные об ископаемых остатках многоклеточных животных, найденных в Канаде и в Китае в породах, имеющих такой геологический возраст и отделенных от пород с остатками здиакарских организмов слоями ледниковых отложений).
Таким образом, в позднем протерозое (600-650 млн лет назад) уже существовали такие группы многоклеточных животных, как губки, кишечнополостные, плоские и кольчатые черви и даже, возможно, предки членистоногих. Судя по достигнутому уровню организации, можно предполагать, что к этому времени обособились также эволюционные стволы нитчатых червей (тип Nemathelminthes), предков моллюсков и предков вторичноротых животных - олигомерных червей.
Докембрийский филогенез Metazoa можно гипотетически представить следующим образом (рис. 32). От колониальных жгутиковых (по мнению ряда авторов - от гетеротрофных форм, принадлежавших к отряду Protomonadida) путем дифференциации и интеграции колонии, с миграцией внутрь колонии клеток фагоцитобласта на заднем полюсе тела, возникли первые многоклеточные животные, организация которых соответствовала фа-гоцителле (по И. И. Мечникову). Мало изменившимися потомками этих древнейших многоклеточных являются современные Phagocytellozoa (Trichoplax adhaerens). Примитивные многоклеточные были свободноплавающими (за счет работы мерцательного эпителия - кинобласта) животными, питавшимися различными микроорганизмами - простейшими и одноклеточными водорослями.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 810; Нарушение авторских прав?; Мы поможем в написании вашей работы!