КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Часть II. Микроэволюция и видообразование 5 страница
|
|
|
|
ГЛАВА 5. РАЗВИТИЕ ПРИСПОСОБЛЕНИЙ
В классическом дарвинизме процесс видообразования рассматривается как одно из следствий контролируемого естественным отбором развития приспособлений организмов к изменяющимся условиям внешней среды и внутривидовой конкуренции, приводящей к дивергенции дочерних видов. В предшествующих главах мы видели, что видообразование и развитие приспособлений представляют собой хотя и взаимосвязанные, но самостоятельные процессы. Это доказывается возможностью обособления новых филетических линий без существенного влияния отбора - при действии дрейфа генов в малых изолированных популяциях, а также путем гибридогенеза, стасигенеза, полиплоидизации. Вместе с тем эволюция в целом имеет отчетливо приспособительный характер, который обеспечивается действием естественного отбора.
Термин "приспособление" (или адаптация) применяется в современной науке в разных значениях. В широком смысле адоптациями являются любые особенности строения, физиологии, поведения и онтогенеза данного вида, которые обеспечивают (вместе с другими особенностями) возможность специфического образа жизни в определенных условиях внешней среды или определенный способ использования конкретных ресурсов внешней среды. Так, для хищных птиц приспособлениями к хищничеству являются характерная форма клюва и когтей, определенный набор пищеварительных ферментов, необходимых для эффективного переваривания животной пищи, и соответствующие формы поведения, используемые при охоте на различных животных (птиц, млекопитающих, рептилий и др.). Иногда адаптацией называют также процесс развития приспособленности организмов.
Единственный фактор, придающий эволюционным изменениям организмов приспособительный характер, - естественный отбор - непосредственно действует в видовых популяциях, т.е. на уровне микроэволюции. Однако его кумулятивное (накапливающее) действие в полной мере проявляется лишь в процессе смены многих поколений на больших отрезках филетических линий, т.е. в масштабе макроэволюции. Именно на этом уровне из част-Ньк адаптации (незначительных изменений фенотипа, приспосабливающих организмы данного вида к особенностям условий его обитания) формируются адаптации общего значения - сложите морфофункциональные комплексы, обеспечивающие жизнедеятельность организмов в широком диапазоне условий. Частные адаптации специфичны для отдельных видов, тогда как адаптации общего значения характеризуют крупные таксоны (отряды, классы и т.п.) и присущи всем видам, входящим в их состав. Например, летательный аппарат птиц, включающий скелет и мускулатуру передних конечностей, а также перья, образующие несущую поверхность крыла, представляют адаптацию общего значения, тогда как особенности строения этих структур у отдельных видов являются частными адаптациями.
Сложные приспособления и проблема их развития
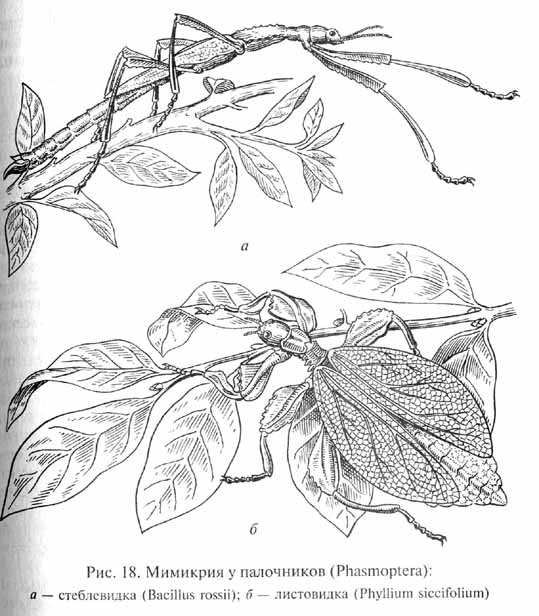
Каждый вид организмов обладает теми или иными приспособлениями, позволяющими ему существовать в определенных условиях среды. Наличие приспособлений придает строению и жизнедеятельности организмов черты целесообразности, которая всегда обращала на себя внимание естествоиспытателей. Невольно поражают воображение сложность и совершенство таких адаптации как, например, взаимные приспособления цветковых растений и опыляющих их насекомых. Сюда относятся характерные особенности строения и окраски венчика цветов и соответствующие особенности строения челюстного аппарата и конечностей насекомых-опылителей, а также черты их поведения. Поразительны явления мимикрии, или подражательного сходства, когда один из видов имитирует особенности внешнего строения и окраски другого, "маскируясь" под несъедобный или ядовитый для хищника объект: "подражание" листу в форме и окраске крыльев или имитация стебля растений у насекомых-палочников (Phasmoptera) (рис. 18), "подражание" неядовитых видов ядовитым или хорошо защищенным (например, уже упоминавшееся выше сходство самок бабочки-парусника с несъедобными бабочками-данаидами, см. рис. 5) и т.п.
Теория Ч.Дарвина впервые дала научное решение проблемы происхождения адаптации как неизбежного результата естественного отбора.
На заре мутационизма многие ученые склонялись к мысли, что новые приспособления возникают сразу готовыми - как случайный результат мутаций крупного масштаба. Подобные воззрения позднее развивал Р. Гольдшмидт, считавший, что крупные "системные мутации" резко изменяют конституцию организма, приводя к появлению "многообещающих уродов" (hopeful monsters), которые могут случайно оказаться приспособленными к новым условиям существования. Эта концепция маловероятна как с генетической, так и с экологической точки зрения. Как уже отмечалось, сколько-нибудь значительные мутации обычно летальны, поскольку они приводят к нарушениям нормальной интеграции организма на том или ином этапе онтогенеза. Маловероятно возникновение посредством крупной мутации такого нового фенотипа, который оказался бы не только жизнеспособным, но и плодовитым, а к тому же априорно приспособленным к каким-либо конкретным условиям существования. В связи с этим Э.Майр справедливо заметил, что организмы, которые возникнут в результате крупных мутаций, скорее окажутся "безнадежными уродами" (hopeless monsters), чем "многообещающими", по Р.Гольдшмидту.
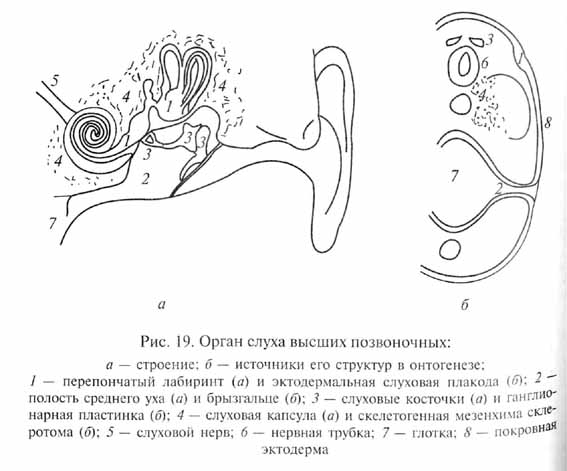
Но наиболее важным возражением против существенной роли крупных мутаций в возникновении новых типов организации является невозможность объяснить этим путем формирование сложных комплексных адаптаций, образованных разнообразными компонентами, развитие которых кодируется разными генетическими системами и обеспечивается независимыми цепями морфо-генетических процессов в онтогенезе. Именно таковы многие адаптации общего значения, составляющие основной план организации высших таксонов. Например, слуховой анализатор у высших наземных позвоночных (амниот) включает структуры, имеющие разное эволюционное происхождение и формирующиеся в онтогенезе разным путем из различных зачатков (рис. 19). Так, внутреннее ухо развивается из слуховой плакоды (утолщение зародышевой эктодермы); полость среднего уха - из зачатка рудиментарной жаберной щели (брызгальца); слуховые косточки среднего уха (звукопроводящий аппарат) - из элементов подъязычной и челюстной висцеральных дуг, возникающих, в свою очередь, из материала так называемой ганглионарной пластинки эктодермального происхождения; скелет слуховой капсулы - из скелетогенной мезодермы головы; наконец, слуховой нерв и мозговые центры слухового анализатора формируются из невральной эктодермы. Совершенно ясно, что невозможно возникновение всей этой сложной, но целостной системы как путем случайного подбора различных мутаций (например, при дрейфе генов), так и посредством одной крупной "системной" мутации. Несомненно, естественный отбор играет решающую роль в накоплении мутаций и интеграции их фенотипических проявлений, обусловливая развитие приспособлений организмов.
Однако в этой проблеме имеется ряд сложных вопросов, и некоторые из них отчетливо представлял уже Ч.Дарвин. Первая трудность связана с тем, как отбор может контролировать начальные этапы развития нового приспособления, когда новая структура (или функция) еще столь слабо выражена, что ее небольшие изменения не могут принести организму каких-либо заметных выгод. Пытаясь разрешить эту проблему, Ч.Дарвин в некоторых случаях даже пытался привлечь "унаследование последствий употребления и неупотребления органов" (например, для объяснения развития асимметричного расположения глаз у камбал или развития цепкого хвоста у южноамериканских обезьян). Очевидно, такое объяснение было шагом назад по отношению к теории естественного отбора и не может быть признано удовлетворительным.
Особенно трудно объяснить действие отбора на первых этапах развития таких сложных органов, которые вообще могут эффективно функционировать только при достаточно высокой степени своего развития. Таковы, например, уже упоминавшиеся нами сложные адаптации - летательный аппарат птиц, звукопроводящая система среднего уха наземных позвоночных, плавательный пузырь костистых рыб, грудная клетка амниот в роли легочного воздушного насоса и т.п. Трудно понять, какую приспособительную ценность для организма могут иметь первые зачатки подобных сложных органов и как на них будет действовать отбор, поскольку естественный отбор не может "предвидеть" будущего развития данного признака и его будущего значения для организма.
Развитие приспособлений на основе преадаптаций
Путь к решению этой проблемы помогает найти концепция морфофункциональных преадаптаций. Сам термин "преадаптация" принадлежит французскому ученому Л. Кено. Кено понимал под преадаптациями такие особенности организмов, которые возникают как случайные вариации, первоначально безразличные для организма (т.е. не приносящие ему ни вреда, ни заметной пользы), нo после изменения условий существования или образа жизни Данного вида оказываются готовыми приспособлениями, полезными в новых условиях. Очевидно, эти представления Кено о развитии приспособлений из случайных преадаптаций без участия естественного отбора примыкают к воззрениям ранних генетиков-мутационистов (см. с. 44). Однако в концепции Кено было все же рациональное зерно, заключавшееся в предположении, что Сложные приспособления к каким-либо конкретным условиям Могут первоначально развиваться в совершенно иной среде обитания и выполнять другую роль, и лишь позднее - в новых условиях - принять на себя новые, прежде им не свойственные функции. Кено справедливо указывал, что освоение новой среды обитания любым видом организмов возможно только при наличии у него таких особенностей строения, физиологии и поведения, которые дают возможность выжить в новых условиях, но возникнуть эти особенности могли только до начала освоения нового образа жизни, т. е. в прежней среде обитания. Такие особенности и являются преадаптивными.
Кено обратил внимание на то, что, в сущности, уже Дарвин подошел к идее преадаптации, анализируя в "Происхождении видов" развитие некоторых приспособлений. Так, швы между костями черепа млекопитающих облегчают прохождение головы детеныша при родах, однако наличие таких сочленений между костями черепа связано совсем не с живорождением, и возникли эти соединения костей в черепе древних позвоночных задолго до появления живорождения. "Лазающие пальмы" Малайского архипелага имеют приспособления в виде крючков, позволяющие им удерживаться на ветках и стволах деревьев, взбираясь по ним вверх. Но точно такие же крючки существуют и у многих нелазающих родственных видов растений, которым они служат как защитные приспособления.
За время своего существования концепция преадаптации претерпела значительные изменения, и ныне ее положения существенно отличаются от взглядов Л. Кено. В первую очередь это относится к отказу от наивных представлений, что преадаптации являются уже готовыми приспособлениями, возникающими во всей их сложности чисто случайно, без участия естественного отбора.
Концепция преадаптации охватывает самые различные процессы и явления, связанные с развитием у организмов новых приспособлений и принятием органами новых функций. Эти процессы и явления могут относиться к особенностям строения, физиологии, индивидуального развития, поведения и образа жизни организмов. В наиболее общем смысле преадаптациями называют такие свойства организмов, которые потенциально имеют приспособительную ценность для еще не осуществленных форм взаимодействий организма и среды или для новых способов функционирования отдельных органов. Так же как и в отношении термина "адаптация", преадаптацией называют и состояние организма (или органа), и процесс развития преадаптивного состояния. Процесс усовершенствования уже существующих приспособлений к уже освоенной среде обитания (или - более узко - процесс совершенствования прежних способов функционирования отдельных органов) называют постадаптацией.
Современная концепция преадаптации включает два основных аспекта - генетико-экологический и морфо-функциональный. Первый из них стоит ближе ко взглядам Л. Кено.
При генетико-экологическом подходе в качестве преадаптивных рассматриваются такие нейтральные и "слабовредные" особенности фенотипа, которые поддерживаются мутационным процессом, эффектом плейотропии, коррелятивными взаимосвязями в организме и тому подобными механизмами и оказываются адаптивными при изменении условий внешней среды. В сущности, ге-нетико-экологическая концепция преадаптации сводится к представлениям о возможности использования видом первоначально неприспособительных признаков (возникших на основе случайных мутаций) для развития адаптации при изменениях направления отбора. Сами по себе эти представления не вызывают возражений, но, в сущности, этот подход является малопродуктивным, так как конкретный анализ развития преадаптивных признаков в этом аспекте отсутствует. Кроме того, генетико-экологическая концепция преадаптации выпускает из поля зрения творческую роль естественного отбора, за что она нередко подвергалась критике. (Отметим, что взгляды сторонников генетико-экологической концепции преадаптации или даже взгляды Л.Кено и других ранних мутационистов часто критикуются в качестве "теории преадаптации". которая к этим взглядам отнюдь не сводится).
Морфофункциональные преадаптации и смена функций
В отличие от генетико-экологической концепции, рассматривающей возникновение преадаптивного состояния на основе случайного подбора мутаций, морфофункциональная концепция, разработанная Д. Симпсоном и В. Боком, подчеркивает развитие преадаптации на базе прежних приспособлений, под непосредственным и непрекращающимся контролем естественного отбора. Усовершенствование приспособлений к существующим условиям может оказаться преадаптацией к новым условиям среды.
Морфофункциональная концепция преадаптации позволяет разрешить ту проблему, о которой мы упоминали в начале этой главы, - развитие сложных приспособлений, способных эффективно функционировать лишь будучи вполне сформированными. Такие приспособления (подобные челюстному аппарату, среднему уху, плавательному пузырю и др.) формируются в эволюции из существовавших прежде органов, имевших иное приспособительное значение. Соответствующие органы сначала приобретали новую функцию (дополнительную к их прежней основной Функции), а затем, если новая функция оказывалась более важной, происходила смена функций органа. Принцип смены функций органов в эволюции был сформулирован А.Дорном в 1875г. Такой путь филогенетических изменений возможен лишь в тощ случае, если имеется орган, подготовленный своей предшествующей эволюцией к принятию новой, прежде не свойственной ему функции, т.е. орган, находящийся в преадаптивном состоянии. Морфофункциональная концепция преадаптации анализирует эволюционный механизм смены функций органов.
В этом аспекте преадаптивное состояние организма (или отдельного органа) возникает не как непосредственный результат случайных мутаций, а на основе предшествующей приспособительной эволюции. Естественный отбор постепенно совершенствует уже существующие приспособления организмов, благоприятствуя тем вариантам, которые обеспечивают более эффективное и экономичное функционирование различных органов. В некоторых случаях при этом может возникнуть как случайный, побочный эффект приспособительной эволюции преадаптивное состояние органа, когда последний получает возможность принять новую функцию в дополнение к его прежней функции. При этом орган получает и новое приспособительное значение (новую адаптивную роль). Если новая адаптивная роль достаточно важна для организма, дальнейшая эволюция данного органа будет направлена отбором по пути совершенствования этого нового приспособления (рис. 20).
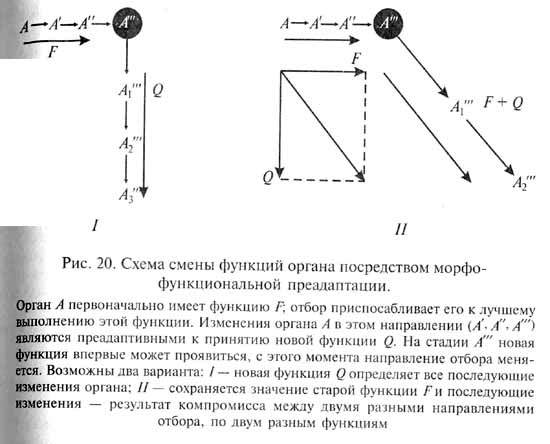
Эволюционный момент, когда орган принимает новую функцию и вследствие этого получает новую адаптивную роль, В.Бок назвал преадаптационным порогом. Прежние функция и адаптивная роль органа могут сохраниться и после принятия им новых. Тогда дальнейшие эволюционные изменения органа определяются компромиссом между двумя направлениями отбора, контролирующими старую и новую адаптации.
Важной особенностью механизма морфофункциональной преадаптации является его пороговый характер: после достижения преадаптационного порога при высокой приспособительной ценности новой адаптивной роли направление эволюционных перестроек данного органа может резко измениться. Одновременно открываются новые возможности в использовании ресурсов внешней среды, что создает предпосылки для интенсивного видообразования, приводящего к возникновению разнообразных вариантов нового конструктивного типа.
Поскольку орган подготовлен к принятию новой адаптивной роли преобразованиями, направленными на усовершенствование прежнего приспособления, т.е. постадаптациями по прежней функции, можно говорить, что преадаптация органа к новой адаптивной роли развивается на основе постадаптаций к прежней.
Потребность в новом приспособлении может существовать у организмов задолго до того, как возникает подходящая преадаптация. Нужно подчеркнуть, что такая потребность далеко не всегда реализуется в эволюции. Поскольку преадаптация возникает как случайный побочный эффект приспособительных эволюционных изменений органа - очевидно, что гораздо чаще должно иметь место как раз отсутствие требуемых преадаптации, чем их развитие. Например, отсутствие предпосылок для вселения вида в новую среду обитания в целом гораздо более обычно, чем наличие требуемых для этого преадаптации.
Целый ряд поразительных примеров смены функций, иллюстрирующих разные формы морфофункциональных преадаптаций, демонстрируют эволюционные преобразования висцерального скелета позвоночных животных. У предков позвоночных (протокраниот, по А.Н.Северцову) висцеральный скелет, вероятно, состоял из серии цельных (нерасчлененных на отделы) хрящевых колец - жаберных дуг, охватывавших передний отдел пищеварительной трубки (рис. 21) и препятствовавших спадению ее стенок. При активизации образа жизни для этих животных стала необходимой интенсификация дыхания. Последняя была достигнута посредством усиления тока воды через жаберную область с развитием здесь нагнетательно-всасывающего насоса. У бесчелюстных позвоночных (предков современного класса круглоротых - Cyclostomata) жаберные дуги потеряли свою первоначальную независимость, соединившись в единую жаберную решетку продольными хрящевыми балками. При вентиляции жабер жаберная решетка, образованная гибкими хрящами, сжимается (при сокращении специальных мышц) и затем расправляется (за счет эластичности хрящевых жаберных дуг) как целое. Этот способ вентиляции жаберной области был, вероятно, связан с развитием энтодермальных жабер, расположенных внутри от жаберных дуг.
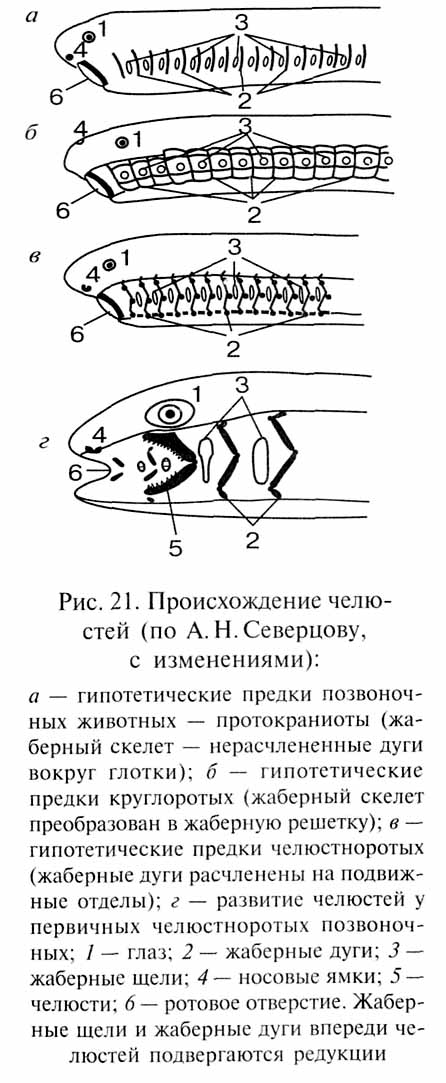
Совершенно иным путем происходила интенсификация обмена воды в жаберной области у предков челюстноротых позвоночных, у которых жабры имеют эктодермальное происхождение и располагаются снаружи от висцерального скелета (см. рис. 21). У этих животных сохранилась самостоятельность отдельных жаберных дуг, и каждая из них расчленилась на несколько подвижных отделов. Сокращение жаберных мышц складывает каждую жаберную дугу как гармошку, энергично сжимая жаберную область. Этот путь эволюции оказался преадаптивным для возникновения челюстей. Передние висцеральные дуги при наличии сильных мышц, сжимающих "гармошку", получили возможность способствовать удерживанию добычи. Подчеркнем, что эта новая адаптивная роль была приобретена на основе преадаптации, связанной с усовершенствованием прежней (дыхательной) роли висцеральных дуг. Новая адаптивная роль (захват и удерживание добычи) оказалась столь важной, что прежняя роль (участие в дыхательных движениях) у передней жаберной дуги (челюстной) отступила на второй план, а затем была утрачена (рудиментарная жабра челюстной дуги сохраняется у всех рыб в виде так называемой ложножабры, теряющей непосредственную связь с челюстной дугой и находящейся у костных рыб на внутренней поверхности жаберной крышки).
Так жаберная дуга превратилась в челюсти - орган схватывания и удерживания добычи. Наличие челюстей стало одним из важнейших преимуществ, способствовавших победе челюстноротых позвоночных над бесчелюстными в борьбе за существование. Среди современных позвоночных челюстноротые составляют примерно 99,8% от общего числа видов, а круглоротые - лишь 0,2 %. Возможно, челюсти не могли развиться у предков круглоротых из-за объединения жаберных дуг в единую жаберную решетку. В этом случае мы наблюдаем как раз такую ситуацию, когда в эволюции большой группы животных (бесчелюстные позвоночные) не была реализована биологическая потребность в развитии челюстей - важного органа добывания пищи, защиты и нападения, так как не возникло необходимой преадаптации.
После прохождения преадаптационного порога в эволюционных изменениях челюстной дуги у челюстноротых позвоночных начался период постадаптаций, усовершенствовавших приспособления к схватыванию и удерживанию добычи. Эти постадаптации привели к возникновению задней опоры челюстной дуги в области челюстного сустава на следующую сзади висцеральную дугу (подъязычную), верхний элемент которой (гиомандибулярный) стал подвеском челюстей. Однако подъязычная дуга сохранила и свою прежнюю роль - участие в вентиляции жаберной области, и у хрящевых рыб несет даже жаберные лепестки. Очевидно, здесь мы имеем пример эволюционного компромисса между двумя направлениями отбора, одно из которых требует сохранения и усовершенствования прежней дыхательной функции подъязычной Дуги, другое - развития новой опорной функции. Оба эти направления отбора контролировали дальнейшие эволюционные преобразования подъязычной дуги у рыб, с одной стороны, обусловив развитие механизма жаберной крышки у костных рыб, позволившего еше более интенсифицировать вентиляцию жаберной области, с другой -- вызвав появление различных механизмов Подвески челюстей к черепу, связанных с разными способами Добывания пищи (амфистилия, гиостилия и др.).
Рассмотренное в этом примере преадаптивное состояние органа возникло в результате усовершенствования его прежней функции; такую форму преадаптации можно назвать собственно (в узком смысле) морфофункциональной. В других случаях преадаптивное состояние может развиваться в процессе редукции органа после утраты его прежней адаптивной роли - редукционная преадаптация. Наконец, третью категорию составляют онтогенетические преадаптации, возникающие на основе эмбрионального или личиночного состояния некоторых органов, сохраняющегося во взрослом организме, в результате задержки онтогенеза на соответствующих стадиях.
Явление редукционной преадаптации можно проиллюстрировать процессом эволюционного развития звукопередающей системы среднего уха у наземных позвоночных-тетрапод. У предков земноводных возник аутостилический череп, в котором гиомандибулярный элемент подъязычной дуги утратил прежнюю роль подвеска челюстей; кроме того, редукция жаберного дыхания сопровождалась прекращением деятельности вентиляционного механизма жаберной крышки, которая также редуцировалась. Потеряв обе свои прежние функции, гиомандибулярный элемент также подвергся некоторой редукции. Этот элемент располагается в черепе сбоку от слуховой капсулы, его отростки упираются в стенку последней и в накладные кости височной области черепа, под которыми здесь располагается полость - рудимент жаберной щели между челюстной и подъязычной дугами (брызгальце). В воздушной среде эта полость оказывается заполненной воздухом; ее наружная стенка после редукции жаберной крышки становится относительно тонкой и, вероятно, могла вибрировать в ответ на колебания окружающего воздуха, как барабанная перепонка. Гиомандибуляре, упиравшееся в эту перепонку и в стенку слуховой капсулы, передавало трансформированные колебания воздуха ко внутреннему уху. Так гиомандибуляре функционально превратилось в первую слуховую косточку среднего уха - слуховой столбик или стремечко. В дальнейшем постадаптации усовершенствовали звукопередаюшие свойства этого элемента. Вероятно, подобная же редукционная преадаптация произошла и у предков млекопитающих после возникновения вторичного челюстного сустава при последовавшей затем редукции костей первичного челюстного сустава - квадратной и сочленовной, превратившихся в наковальню и молоточек среднего уха.
Примером эволюционного использования онтогенетических преадаптации может служить сохранение эмбрионального хрящевого скелета во взрослом состоянии, характерное для некоторых групп позвоночных (хрящевые рыбы, круглоротые, отчасти бесхвостые и хвостатые земноводные). В приспособительном отношении это было, вероятно, связано с необходимостью облегчения скелета (см. ч. III) при активизации образа жизни или при выходе на сушу.
Для освоения новой среды обитания, существенно отличающейся от прежней, организмы должны обладать целым комплексом особенностей строения, физиологии и поведения, позволяющих им существовать в этой новой среде. Такое состояние называется комплексной или конституциональной преадаптацией. Например, древнейшие земноводные, начавшие освоение суши Как среды обитания, обладали таким строением конечностей, органов дыхания и кровообращения и специфическими особенностями образа жизни и поведения, вся совокупность которых была необходима для выхода на сушу из водоемов. Все эти особенности, составившие комплексную преадаптацию, развились как специфические приспособления предков земноводных - кистеперых рыб - к жизни в пресных водоемах тропического пояса, в которых частым явлением было значительное уменьшение растворенного в воде кислорода (см. ч. III). В комплексной преадаптации, в сущности, объединяются морфофункциональный и генетико-экологический аспекты преадаптадии. Как указал Г.Оше, комплекс уже сложившихся морфофункциональных преадаптации для своей реализации должен быть завершен некоторыми деталями, которые могут контролироваться немногими ключевыми мутациями. Например, эти мутации могут вызвать такие изменения физиологии или поведения, которые позволят данному виду начать освоение новой среды обитания. Это и преобразует комплекс преадаптации в новое приспособление.
В связи с этим упомянем еще один интересный пример преадаптации, замечательный тем, что он реализовался не в далеком прошлом, как упоминавшиеся выше, а в наше время. Такие особенности строения попугаев, как крупный острый клюв с изогнутым вниз надклювьем (рис. 22), крепкие острые когти, мощная мускулатура челюстного аппарата, шеи и конечностей, выглядят хорошо приспособленными для хищного способа питания. Однако все эти признаки, несомненно, сформировались как приспособления к лазанию По деревьям: при лазании клюв также используется как хватательный орган, своего Рода "пятая конечность", тем более что надклювье подвижно сочленяется с черепом, и клюв как целое может активно подгибаться вниз или отгибаться вверх (что нетрудно заметить, наблюдая за лазанием крупных попугаев по решетке клетки). Подавляющее большинство попугаев - мирные птицы, питаЮщИеся растительной пищей, главным образом различными плодами и семенами.

Однако известен один вид попугаев, который действительно использовал указанные признаки при освоении хищного образа жизни. Попугай кеа (Nestor notabilis) (см. рис. 22), обитающий в горных районах южного острова Новой Зеландии, питается обычно различной растительной пищей (фрукты, нектар цветов), а также мелкими беспозвоночными животными (черви, насекомые) которых добывает из-под камней. Однако начиная с 1868 г. были отмечены случаи нападения кеа на овец, пригоняемых на горные пастбища. Хищнические наклонности проявляют отдельные особи попугаев: они расклевывают на спине овец огромные раны (до 10 см), от которых жертва быстро погибает. Мясо погибших овец поедают не только птицы-убийцы, но и другие кеа. Поскольку овцеводство появилось в Новой Зеландии после прихода туда европейцев (до этого на острове не было других млекопитающих, кроме крыс и летучих мышей), очевидно, что переход попугаев кеа к хищничеству произошел, можно сказать, на наших глазах. При этом строение клюва и когтей попугаев не претерпело никаких изменений - их использование для новой роли было преадаптировано прежними приспособлениями, облегчавшими попугаям лазание по деревьям и питание плодами растений. Попугаи, нападающие на овец, ничем не отличаются от других кеа. Возможно такое изменение поведения обусловлено возникновением соответствующей мутации, позволившей использовать по-новому существовавшие прежде структуры.
Необходимо подчеркнуть, что путь преадаптации - широко распространенный, но не единственно возможный способ развития новых приспособлений. Некоторые структуры, особенности физиологии и поведения могут с самого начала формироваться как "постадаптации" (здесь термин чисто условный, поскольку отсутствует период преадаптации). Условием для этого является такая приспособительная ценность уже первых зачатков новой структуры, которая достаточна для возникновения положительного давления естественного отбора, благоприятствующего новому признаку. Примером сложного органа, возникшего таким путем, является глаз, как это убедительно показал Ч.Дарвин.
Относительность приспособленности и инадаптивная эволюция
При высокой степени совершенства многих адаптации еще Ч.Дарвин подчеркивал относительность приспособленности и воз можность в большинстве случаев появления лучше приспособленных организмов, чем уже существующие. При отсутствии конкуренции приспособления совершенствуются отбором не максимально возможного, а до некоторого удовлетворительного уровня, после чего интенсивность отбора существенно падает. Достигаемая в каждом конкретном случае степень совершенства любого приспособления определяется, с одной стороны, особенностями организации данного вида, с другой же - интенсивностью отбора, в свою очередь зависящей от степени соответствия видовых приспособлений существующим внешним условиям. Кроме того, развитие разных адаптации у данного вида организмов может в какой-то мере противоречить друг другу, ставя различного рода взаимные ограничения. В подобных ситуациях отбор благоприятствует формированию некоторого оптимального соотношения между разными адаптациями, а не достижению максимального совершенства той или другой из них (принцип адаптивного компромисса, по К.Гансу).
Относительность приспособленности организмов ярко проявляется в многочисленных случаях неудачного решения тех или иных эволюционных проблем в филогенезе различных групп организмов. Такой "неудачный выбор" направления эволюционных преобразований, который может раньше или позже привести к эволюционному тупику, к назреванию неразрешимых или трудноразрешимых внутренних противоречий, был назван В.О.Ковалевским в 70-е гг. XIX в. инадаптивной эволюцией.
В.О.Ковалевский проанализировал это явление на примере филогенетических преобразований конечностей некоторых парнокопытных млекопитающих. В эволюции различных групп копытных в связи с приспособлением к быстрому бегу по относительно твердой почве происходила редукция боковых пальцев. Эта редукция у разных филетических линий ископаемых парнокопытных осуществлялась по-разному. У некоторых форм (Dicotyles, Gelocus, Sus) редукция пальцев происходила вместе с соответствующими перестройками в запястье и предплюсне, так что вес животного передавался на усиливающиеся средние пальцы (третий и четвертый), а конечность в целом сохраняла прочность и устойчивость (адаптивный путь изменений). У других же парнокопытных (Anoplotherium, Anthracotherium, Entelo-don, Hyopotamus, Xiphodon) редукция боковых пальцев опережала перестройку запястья и предплюсны, сохранявших более примитивное строение (рис. 23). В этом случае возникала механически несовершенная конструкция конечности, которая не обеспечивала равномерного распределения нагрузок в скелете при дальнейшей редукции боковых пальцев (инадаптивный путь). Преобразования конечностей по инадаптивному пути происходили в целом быстрее, чем более сложные гармонические перестройки по адаптивному пути. В результате этого инадаптивный путь изменений способствовал более раннему расцвету соответствующих филетических линий, но в дальнейшем при конкуренции с представителями более медленно, но гармонично развивавшихся линий инадаптивные формы вымерли.
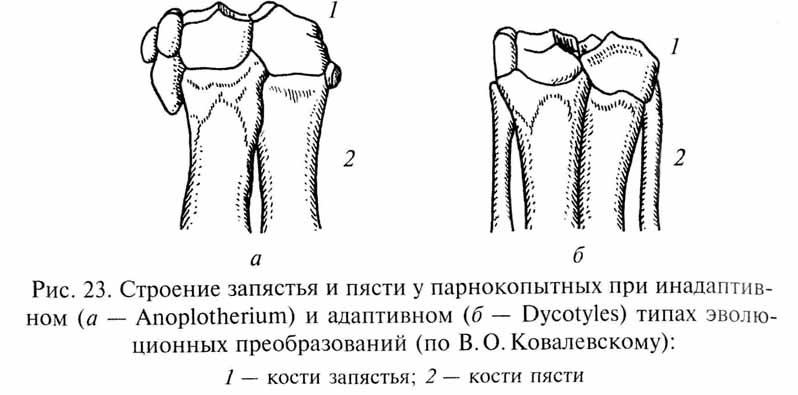
При анализе подобных ситуаций неизбежно возникает вопрос: как объяснить развитие инадаптивных изменений? Не противоречит ли "инадаптивная эволюция" представлениям о развитии приспособлений под контролем естественного отбора? Однако противоречия здесь нет. Дело в том, что на первых этапах эволюционных преобразований многих органов (в данном случае - в процессе редукции боковых лучей конечностей у парнокопытных) и адаптивные, и инадаптивные варианты изменений могут быть примерно в равной степени полезны для вида. Естественный отбор благоприятствует любому изменению, повышающему приспособленность организмов, вне зависимости от того, как пойдет развитие этого признака в дальнейшем. Строго говоря, термин В.О.Ковалевского "инадаптивная", т.е. неприспособительная, эволюция неудачен, так как отбираются только приспособительные (адаптивные) изменения организмов. Однако в ходе дальнейшей эволюции обнаруживается преимущество одного пути эволюционных преобразований перед другими. Механическое несовершенство конечностей "инадаптивных" форм копытных выявилось лишь при достаточно далеко зашедшей редукции боковых пальцев, а в начале этого процесса указанные формы, вероятно, ничем не уступали представителям "адаптивных линий".
Таким образом, можно назвать инадаптивным такой путь филогенетических изменений, при котором формирующееся приспособление заключает в себе предпосылки для возникновения тех или иных внутренних противоречий, которые нарастают мере развития приспособления, приводя в конечном итоге к эволюционному тупику. Как показал В.О.Ковалевский, на начальных стадиях развития инадаптивные изменения могут даже и определенные преимущества перед более медленно протекающими гармоническими преобразованиями - именно благодаря быстроте развития инадаптивных вариантов. Гармонические преобразования требуют большего числа эволюционных "проб и ошибок" и соответственно большего времени для своего осуществления. Вероятно, можно сказать, что быстрота эволюционных преобразований нередко оказывается обратно пропорциональной степени их совершенства.
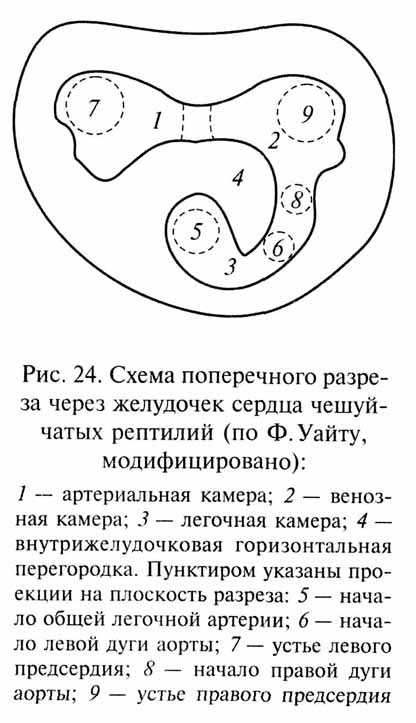
Инадаптивный путь эволюции не обязательно ведет к вымиранию - возникшие противоречия могут быть различными способами компенсированы. Рассмотрим, например, эволюционные преобразования сердца у рептилий завропсидного ствола (черепахи, клювоголовые, ящерицы, змеи, крокодилы). У предков завропсидных пресмыкающихся в желудочке сердца возникла горизонтальная перегородка, разделившая его на спинную и брюшную камеры и способствовавшая распределению артериальной крови в дуги аорты, а венозной - в легочную артерию (рис. 24). Однако полное разделение желудочка этой перегородкой, которое совершенно прекратило бы смешивание артериальной и венозной крови, невозможно - при этом легочная артерия, начинающаяся от брюшной камеры желудочка, оказалась бы отрезанной от обоих предсердий, открывающихся в спинную камеру. В результате у большинства пресмыкающихся перегородка в желудочке сердца остается неполной и сохраняется смешивание артериальной и венозной крови в сердце, что препятствует интенсификации метаболизма до уровня, необходимого для достижения гомойотермии (независимости температуры тела от температуры внешней среды). У предков крокодилов и птиц первичная горизонтальная перегородка в желудочке сердца редуцировалась и вместо нее возникла новая - вертикальная перегородка, полностью разделившая желудочек на левую (артериальную) и правую венозную) камеры. После редукции у предков птиц левой дуги начинавшейся от правой камеры желудочка, полностью прекратилось смешивание артериальной и венозной крови, что стало одной из важных предпосылок для развития у птиц гомойотермии. У предков млекопитающих в желудочке сердца сразу возникла вертикальная перегородка, которая, пока она была неполной, вероятно, не давала этим животным каких-либо преимуществ в разделении артериальной и венозной крови по сравнению с "типичными" пресмыкающимися - но этот путь эволюционных преобразований сердца позволил в дальнейшем разделить желудочек полностью без сложных дополнительных перестроек. Можно полагать, что предки завропсидных рептилий пошли по "инадаптивному" пути в разделении желудочка сердца по сравнению с более удачным ("адаптивным") решением этой проблемы у терапсидных рептилий - предков млекопитающих.
Инадаптивные изменения неизбежно должны быть обычным явлением в филогенезе различных групп организмов, ибо единственный фактор эволюции, обусловливающий развитие приспособлений, - естественный отбор - действует, так сказать, "в настоящем времени" и не может предвидеть эволюционных последствий преобразований, благоприятных в данный исторический момент и при данных условиях, но могущих при их дальнейшем развитии привести к эволюционному тупику и вымиранию.
Гиперадаптивность
На фоне обшей относительности совершенства приспособлений, наглядно проявляющейся в явлениях инадаптивной эволюции, особенно контрастно выглядит так называемая гиперадаптивность или конструктивная избыточность многих биологических систем, представляющая собой более или менее значительное (в некоторых случаях многократное) превышение их адаптивных возможностей по отношению к обычным условиям их функционирования. Это относится, например, к прочности костей на разрыв и сжатие (у млекопитающих она близка к прочности стали), к силе сокращения мышц (для одной мышцы лягушки она может примерно в 5 раз превышать вес всего тела) и т.п. Поразительную гиперадаптацию к действию низких температур проявляют в состоянии анабиоза споры ряда бактерий, а также некоторые микроскопические многоклеточные животные - коловратки (Rotatoria - класс типа нитчатых червей) и тихоходки (Tardigrada -- группа, близкая к кольчатым червям и членистоногим), которые могут выдержать температуру жидкого гелия (-270°С), близкую к абсолютному нулю.
Проявлением гиперадаптивности является также наличие в составе некоторых органов такого количества структурных элементов, которое во много раз превышает необходимое для их нормального функционирования при обычных условиях. Например, в женском яичнике закладывается от 40 до 200 тыс. яйцевых фолликулов, из которых овулирует в течение всей жизни женщины не более 500.
Важность и необходимость гиперадаптивности для выживания вида в изменчивой среде очевидна. В истории любого вида организмов неизбежно встречаются периоды необычных (экстремальных) условий, ставящих под угрозу существование целых популяций. Проявления гиперадаптивности представляют собой "запас прочности" систем организма, позволяющий им выдерживать повышенные функциональные нагрузки в таких экстремальных условиях. Конструктивная избыточность многократно повышает надежность биологических систем и является необходимым условием для выживания организмов, когда давление отбора достигает предельных значений.
Однако механизм возникновения гиперадаптивности является одной из сложных проблем адаптациогенеза. Естественный отбор, действуя лишь "в настоящем времени", может благоприятствовать только таким изменениям организмов, которые обеспечивают их носителям при данных условиях какие-либо преимущества в выживании или оставлении потомства. В то же время подавляющее большинство особей данного вида в течение всей жизни не подвергается воздействию экстремальных условий, при которых обладатели гиперадаптивного состояния получили бы соответствующие преимущества. Следовательно, гиперадаптивные признаки сами по себе имеют чрезвычайно мало шансов быть "замеченными" естественным отбором, и еще менее вероятны накопление и интеграция соответствующих изменений в ряду последовательных поколений. И совсем уже невероятным выглядит прямое закрепление отбором таких гиперадаптивных особенностей, которые обеспечивают сверхустойчивость организма (вплоть до условий, вообще не наблюдающихся ныне в биосфере Земли, например температур, близких к абсолютному нулю).
Чтобы разобраться в этой проблеме, прежде всего отметим, что понятие гиперадаптивности объединяет широкий круг очень разнородных явлений (в этом легко убедиться на приведенных выше примерах). Вероятно, возникновение разных форм конструктивной избыточности определяется различными причинами.
Вообще для нормального функционирования любого органа При самых обычных (средних) условиях полезно, чтобы диапазон его функциональных возможностей был существенно шире обычно используемого. Это позволяет избежать сбоев, вероятных в крайних областях функционального диапазона: краевые зоны неустойчивого функционирования отодвигаются за пределы обычно используемой "рабочей части" этого диапазона. Другими словами, некоторая потенциальная гиперадаптивность полезна уже для нормального функционирования органа в обычных условиях. Поэтому естественный отбор в процессе совершенствования любой адаптации благоприятствует расширению функциональных возможностей органа в гиперадаптивную область.
Дальнейшему усилению гиперадаптивности способствует действие катастрофического отбора (см. с. 108) в экстремальных условиях, раньше или позже неизбежно возникающих в истории любой филетической линии. При действии катастрофического отбора вид может выжить только в том случае, если в его популяциях имеются особи, обладающие соответствующей конструктивной избыточностью по сравнению с "нормой". При крайне неблагоприятных условиях катастрофический отбор элиминирует практически всю прежнюю популяцию, за исключением особей-носителей гиперадаптивного состояния отбираемых признаков. После возврата условий к прежней норме выжившие особи формируют новую популяцию, потомки которой будут сохранять конструктивную избыточность просто как своего рода нейтральные признаки (поскольку в нормальных условиях высокая степень гиперадаптивности отбором не контролируется) до следующей экстремальной ситуации. Очевидно, в длительной истории филетической линии важность гиперадаптаций для выживания вида не вызывает сомнений. Конструктивная избыточность, обеспечивающая запас прочности систем организма и дающая виду шансы на выживание в экстремальных условиях, является результатом действия катастрофического отбора в прошлой истории вида.
Кроме того, как показал П.В.Пучков, многие проявления конструктивной избыточности возникают как побочный результат контролируемого отбором развития обычных адаптации организмов к специфическим условиям их обитания. Так, гиперадаптивная устойчивость яиц жаброногих раков к действию низких температур (до -180°С), вероятно, возникла как побочный результат развития приспособлений, позволяющих этим организмам выживать при длительном пересыхании временных водоемов (благодаря особому химизму яиц и структуре их оболочек яйца жаброногих раков выдерживают высыхание до 9 лет), а также проходить без повреждений сквозь кишечный тракт проглотившего яйцо хищника.
Этот путь развития гиперадаптивных особенностей, очевидно, соответствует рассмотренному выше механизму морфофункиио-нальной преадаптации, обеспечивающему возможность расширения функций органов и приобретения ими новой адаптивной роли. Механизм преадаптации позволяет объяснить наиболее значительные проявления конструктивной избыточности организмов.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 603; Нарушение авторских прав?; Мы поможем в написании вашей работы!