КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Часть II. Микроэволюция и видообразование 1 страница
|
|
|
|
Необходимыми предпосылками эволюционного процесса являются возникновение элементарных изменений аппарата наследственности - мутаций, их распространение и закрепление в генофондах популяций организмов. Направленные изменения генофондов популяций под воздействием различных факторов представляют собой элементарные эволюционные изменения -микроэволюцию. Микроэволюция завершается формированием новых видов организмов (процесс видообразования). Рассмотрение предпосылок, структурных элементов, факторов и механизмов всех этапов микроэволюции является основным содержанием этой части книги.
Чтобы не нарушать последовательность изложения, в нее пришлось включить также ряд проблем, выходящих за рамки собственно микроэволюционных процессов. Это относится прежде всего к различным межвидовым отношениям, которые оказывают значительное влияние на процессы видообразования. Межвидовые отношения реализуются в природных сообществах разных видов организмов -- биоценозах, представляющих собой целостные биологические макросистемы с механизмами авторегуляции. Их преобразования могут иметь уже макроэволюцион-ное значение.
Заключительная глава этой части посвящена развитию приспособлений организмов. Эта проблема также занимает пограничное положение между микро- и макроэволюцией. Формирование приспособлений организмов (адаптациогенез) начинается на уровне микроэволюции, но общий приспособительный характер эволюционного процесса и ряд специфических проблем адаптациогенеза в полной мере проявляются лишь в масштабе макроэволюции.
Таким образом, разделы второй части, посвященные межвидовым отношениям и развитию приспособлений, предваряют переход к проблемам макроэволюции, которым будут посвящены третья и четвертая части книги. Соотношения микро- и макроэволюции как двух уровней системной организации эволюционного процесса и свойства разных категорий биологических систем будут рассмотрены в начале четвертой части.
ГЛАВА 1. ЭЛЕМЕНТАРНЫЕ ПРЕДПОСЫЛКИ ЭВОЛЮЦИОННОГО ПРОЦЕССА: ФОРМЫ ИЗМЕНЧИВОСТИ ОРГАНИЗМОВ
Эволюция основывается прежде всего на возникновении наследственных изменений организмов, которые представляют собой необходимый исходный материал для осуществления эволюционного процесса и, таким образом, являются элементарными предпосылками последнего. Особую роль в приспособлении биологических видов к изменяющимся условиям внешней среды играют и ненаследственные изменения организмов. Для выяснения природы разных форм изменчивости организмов, их соотношений друг с другом и для анализа их эволюционной роли нам необходимо сначала хотя бы кратко остановиться на рассмотрении сущности наследственности.
Современные представления о наследственности организмов
Наследственность организмов, под которой понимается способность передавать от поколения к поколению основные структурные и функциональные свойства, обеспечивающие сходство организации потомков и их родителей, представляет собой одно из фундаментальных качеств живых организмов. Само по себе явление наследственности чрезвычайно давно известно людям, но сущность этого важнейшего свойства организмов стала понятной лишь в середине XX в., когда была доказана роль хромосомной дезоксирибонуклеиновой кислоты (ДНК) в передаче наследственных свойств, а в 1953 г. Дж. Уотсоном и Ф. Криком была расшифрована структура молекулы ДНК.
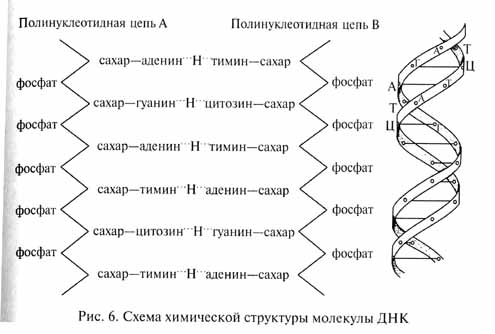
Хромосомы состоят из молекул ДНК, рибонуклеиновой кислоты (РНК) и некоторых типов белков. Основную роль в аппарате наследственности играет ДНК. Согласно модели Уотсона и Крика, молекула ДНК состоит из двух полинуклеотидных цепочек (рис. 6), спирально закрученных вокруг общей оси. Каждый отдельный нуклеотид включает молекулу (точнее, молекулярный остаток) сахара-циклопентозы (в ДНК - дезоксирибоза) и связанные с ней молекулярные остатки фосфорной кислоты и одного из азотистых оснований (пуриновых - аденина и гуанина, или пиримидиновых - тимина и цитозина). Нуклеотиды связаны друг с другом в полинуклеотидную цепочку, а две соседние полинуклеотидные цепочки связаны друг с другом в одну дву-спиральную молекулу ДНК водородными связями между пури-новыми и пиримидиновыми основаниями разных цепочек. При этом возможны только соединения аденин-тимин и гуанин-цитозин. Нуклеотиды с соответствующими парами оснований могут чередоваться в молекуле ДНК в любом порядке. Именно эта последовательность чередования разных пар азотистых оснований молекулы ДНК и представляет собою запись наследственной информации. При этом различные комбинации оснований в последовательных тройках (триплетах) нуклеотидов кодируют разные аминокислоты в молекулах полипептидов, синтезируемых в клетке. Отдельные гены представляют собой участки молекулы ДНК, обладающие определенной биохимической функцией (например, ответственные за синтез определенного типа белковых молекул).
Молекулы ДНК обладают способностью к редупликации, т.е. они могут удваиваться, причем новые молекулы в норме совершенно идентичны старой по строению и расположению всех нуклеотидов. На этом свойстве редупликации ДНК основана точная передача наследственных признаков от поколения к поколению.
Второе важнейшее биологическое свойство молекул ДНК - их способность контролировать синтез белков в клетке, причем, естественно, специфика молекул ДНК у данного вида организмов определяет специфику белкового синтеза у этого вида. Перс-дача информации с ДНК в цитоплазму для последующего синтеза белков на рибосомах осуществляется через посредство РНК -более простой одноцепочной нуклеиновой кислоты, в молекуле которой место тимина занимает другое пиримидиновое основание - урацил. Процесс синтеза молекул РНК на матрице ДНК (так называемая транскрипция) происходит при участии внутриклеточного фермента транскриптазы (или ДНК-зависимой РНК-полимеразы).
Точность редупликации молекул ДНК в процессах клеточного деления (митоза) обеспечивает полную генетическую эквивалентность всех клеток многоклеточного организма. Каждая клетка содержит полный геном, т.е. минимальный полный набор наследственных факторов, включающий 100% генетической информации. Однако в разных типах соматических клеток (например, эпителиальных, нервных, мышечных и т. п.) синтезируются совершенно определенные белки, специфичные для каждого типа клеток. Это обусловлено инактивацией (репрессией) большей части генов в хромосомах соматических клеток хромосомными белками (по одной из гипотез - белками-гистонами). Таким образом, специфичность белкового синтеза в клетках, образующих разные ткани и органы, определяется активностью различных участков хромосом.
Непосредственно кодируют последовательность аминокислот в молекулах полипептидов, синтезируемых на матрицах нуклеиновых кислот, далеко не все гены, а лишь некоторые из них, называемые структурными генами. Другие гены - регуляторы -контролируют процессы считывания наследственной информации со структурных генов, определяя синтез белков, репрессирующих те или иные структурные гены.
Новейшие молекулярно-генетические исследования показали значительную сложность организации генома. Оказалось, что в геномах большинства организмов содержится гораздо больше ДНК, чем необходимо для обеспечения биосинтеза всех белков данного организма. Значительная часть генома представлена так называемой "молчащей ДНК", не транслируемой в белки (трансляцией называется процесс синтеза полипептидных цепей белка по матрице информационной РНК; трансляция происходит на рибосомах). Она включает участки с многократными повторами одних и тех же последовательностей нуклеотидов. Кроме того, у эукариот (т. е. организмов, клетки которых имеют обособленное ядро, см. с. 149) гены обладают своего рода мозаичной структурой: транслируемые при синтезе белков участки (экзоны) чередуются с нетранслируемыми (нитронами).
С другой стороны, в геноме многих организмов были обнаружены так называемые мобильные генетические элементы - фрагменты молекул ДНК, которые могут перемещаться по геному, "встраиваясь" с помощью синтезируемых на их матрицах специальных ферментов в разные места хромосом и вызывая разнообразные наследственные изменения, подавляя или усиливая проявления активности других генов и способствуя повышению мутагенеза (см. ниже). Показано, что у эукариот мобильные генетические элементы составляют не менее 5-10% всего генетического материала.
Мобильные генетические элементы имеют различные размеры (порядка 1000-20000 нуклеотидов) и структуру. Наиболее крупные и сложные из них - плазмиды - способны не только интегрироваться в состав хромосом, образуя так называемые эписомы, но и самостоятельно функционировать в клетке. Полагают, что мобильные генетические элементы могут перемещаться из одной клетки в другую и даже переносить наследственную информацию между разными видами организмов (подробнее см. кн.: Xесин Р.Б. Непостоянство генома. - М., 1984). Подобный так называемый "горизонтальный" (или "латеральный") межвидовой перенос генетической информации, видимо, достаточно широко распространен у прокариот (низшие безъядерные организмы - бактерии, синезеленые водоросли); для эукариот хорошо доказанных данных такого переноса пока нет.
Помимо клеточного ядра специфическая ДНК локализована также в небольшом количестве в митохондриях и пластидах. По-видимому, ДНК этих органоидов обладает такими же основными свойствами, как и ядерная ДНК (способность к редупликации и контролю за синтезом определенных белков через посредство РНК), и представляет собой основу аппарата цитоплазматической наследственности (или плазмотипа), который дополняет основной, ядерный, наследственный аппарат -- генотип. Плазмотип состоит из относительно небольшого (30-300) количества плаз-могенов, кодирующих синтез некоторых белков, важных для функционирования соответствующих органоидов.
ДНК является основной, но, возможно, не единственной формой передачи наследственной информации. Дополнительными источниками информации, передающейся от клетки к клетке, могут быть некоторые надмолекулярные структуры (особенно система клеточных мембран). По мнению К. Маркерта и Г.Уршпрунга, они могут служить матрицами для сборки аналогичных структур из отдельных молекул. Вместе с ДНК митохондрий и пластид эти механизмы передачи наследственной информации входят в состав плазмотипа. В некоторых случаях плазмотип обеспечивает проявление так называемой "материнской наследственности", которая заключается в преобладании у гибридного организма материнских особенностей. Известный пример материнской наследственности - различия мулов (потомство от скрещивания жеребца с ослицей) и лошаков (потомство от скрещивания осла с лошадью). В подобных случаях преобладающее сходство потомства с матерью обеспечивается передачей наследственной информации цитоплазмой яйцеклетки (сперматозоид по сравнению с яйцеклеткой практически лишен цитоплазмы).
В принципе передача наследственных свойств плазмотипом вполне сходна с таковой, осуществляемой генотипом. Очевидно, цитоплазматическая наследственность не имеет никакого отношения к "адекватной соматической индукции" по той же самой причине, по которой фенотипические изменения, как таковые, не передаются следующим поколениям через генотип (см. выше).
Норма реакции, модификационная изменчивость и проблема обратной транскрипции
Необходимо подчеркнуть, что наследственный аппарат сам по себе кодирует только синтез специфических белковых молекул, а отнюдь не признаки целого сформированного фенотипа (т.е. конкретные особенности строения и функционирования тех или иных органов), как это иногда упрощенно представляют. Синтез специфических белков в зародышевых клетках и взаимодействие этих клеток друг с другом определяют общее направление морфогенетических процессов в ходе эмбрионального развития организма. Сущность явлений, лежащих в основе перехода наследственной информации с молекулярного уровня на клеточный в развивающемся организме, во многом еще не ясна. Однако вполне очевидно, что все морфогенетические процессы протекают в разных условиях несколько по-разному. Генотип программирует лишь общее направление морфогенетических процессов, а условия внешней среды оказывают существенное влияние на их конкретную реализацию. В результате каждый ген имеет несколько разных вариантов своего фенотипического проявления, т. е. разных состояний того признака или признаков, которые контролируются этим геном. Всякий конкретный генотип в разных условиях обеспечивает развитие разных фенотипов, т.е. разных совокупностей морфологических, физиологических и поведенческих признаков организма. Все эти фенотипы составляют норму реакции одного и того же генотипа на различные внешние условия. По удачной формулировке М.М.Камшилова, фенотипы наследственно обусловлены, но, как таковые, не наследуются: наследственна лишь норма реакции всякого конкретного генотипа на внешние воздействия.
Поскольку норма реакции обеспечивает формирование тех или иных фенотипов в соответствии с определенными условиями среды, она имеет огромное значение для выживания вида и, следовательно, подвергается действию естественного отбора. Поэтому фенотипические признаки, проявляющиеся у всякого организма в зависимости от условий его сушествования, обычно имеют черты "целесообразности", являясь приспособлениями к данным условиям.
Эти разные фенотипические проявления одних и тех же генов представляют собой изменчивость, но изменчивость, не являющуюся для вида чем-то новым, поскольку все ее варианты запрограммированы в норме реакции генотипа, чтобы обеспечить выживание организмов данного вида в разных условиях. Это и есть модификационная (или фенотипическая) изменчивость, на которой концентрируют внимание ламаркисты, принимающие ее за основу эволюционного процесса. Этой форме изменчивости соответствует определенная изменчивость в теории Дарвина. Из всего сказанного совершенно очевидно, что эта изменчивость хотя нередко и приобретает в результате отбора приспособительный характер, но не может сама по себе привести ни к каким эволюционным преобразованиям.
Если организм подвергается воздействию необычных для данного вида экстремальных условий, возникающие при этом модификации нередко теряют приспособительный характер, получая облик уродства; такие модификации называются морфозами.
Неоламаркисты пытались привлечь для аргументации своих позиций некоторые новые достижения молекулярной генетики, в частности - возможность обратной передачи наследственной информации - с РНК на ДНК. Эта так называемая обратная транскрипция происходит в специфических условиях инфицирования клеток эукариот особыми вирусами (онко-РНК-вирусы), у которых геном образован РНК. При действии вирусного фермента РНК-зависимой ДНК-полимеразы, или ревертазы, на матрице вирусной РНК синтезируется молекула ДНК, которая затем внедряется в геном инфицированной клетки. Сторонники неоламаркистских взглядов полагают, что этот механизм может быть использован Для объяснения адекватной соматической индукции. При этом не обращают внимания на крайнюю специфичность ситуации, при которой происходит обратная транскрипция - вирусная инфекция. Но даже если и допустить подобную возможность, это отнюдь не докажет желаемого для неоламаркистов унаследования "целесообразных" изменений соматических структур, возникших как приспособительные реакции на изменения внешних условий.
Вариации строения и функций соматических структур приобретают приспособительный характер только в результате эволюции под контролем естественного отбора. Все целесообразные приспособительные реакции фенотипа являются модификациями - и никакой механизм передачи наследственной информации не закрепит этих признаков просто потому, что они уже входят в норму реакции данного генотипа, не являясь, в сущности, новыми признаками.
Конечно, изменения внешних условий могут вызвать и такие нарушения в геноме соматических клеток, которые приведут к возникновению качественных новообразований - соматических мутаций. Однако эти изменения, как и все мутации, не являются приспособительными (целесообразными) -- их фенотипические проявления имеют случайный характер по отношению к факторам, вызвавшим их появление. Подробнее наследственную изменчивость мы рассмотрим в следующем разделе этой главы.
Мутационная изменчивость
Эволюционные преобразования организмов основываются на изменениях аппарата наследственности, приводящих к соответствующим перестройкам нормы реакции генотипа; такие изменения получили название мутаций. Мутации могут возникнуть (и неизбежно возникают в определенном проценте случаев) в процессе редупликации генома, предшествующем клеточному делению, или в процессе самого клеточного деления. Точные причины естественно происходящих (спонтанных) мутаций обычно неизвестны, но, очевидно, они кроются в каких-то нарушениях нормального режима сложных процессов редупликации и расхождения хромосом, связанных в конечном итоге с внешними воздействиями на клетку. Возникновение спонтанных мутации связывают, например, с мутагенной активностью некоторых метаболитов (продуктов обмена веществ) или с термическими флюктуаииями атомов в процессах редупликации и т. п.
Размножение и связанная с ним редупликация наследственного аппарата представляют собой важнейшие и неотъемлемые свойства живых организмов. Поскольку эти сложные процессы протекают на фоне постоянных изменений внешней среды и поскольку всякий механизм и всякая структура обладают конечной степенью устойчивости, нарушения процесса редупликации, ведущие к мутациям, совершенно неизбежны. Мутации представляют собой элементарные изменения, являющиеся материалом для действия естественного отбора. Неизбежный результат естественного отбора - эволюционные изменения организмов. Таким образом, как отметил Н.В.Тимофеев-Ресовский, неизбежность эволюции следует из основных свойств живой материи.
Хотя стимулом мутаций являются изменения внешних условий, между этими последними и происходящими под их влиянием мутациями нет адекватности. Это означает, что возникающие мутации сами по себе не являются приспособлениями к изменениям условий, вызвавшим их появление. Аппарат наследственности не обладает и принципиально не может обладать свойством целесообразно перестраиваться в ответ на изменения внешней среды. Это отчетливо проявляется при искусственном мутагенезе, когда внешние воздействия, вызвавшие возникновение мутаций, точно известны. Например, хорошо известно мутагенное действие проникающей радиации, но возникшие под ее влиянием мутации могут затрагивать самые различные особенности организмов (цвет глаз, степень развития и особенности строения крыльев у мух и т. п.), которые в подавляющем большинстве случаев никак не связаны с защитой организма от радиации. Точно так же действуют и другие, более "мягкие" мутагены (различные химические факторы, изменения температуры и др.), причем разные мутагены могут вызывать у данного вида организмов одни и те же мутации.
Этот факт важен в двух отношениях. С одной стороны, он подчеркивает неопределенный, неприспособительный характер мутаций, с другой - многократное появление у разных особей данного вида организмов одних и тех же мутаций. Из этого следует чрезвычайно важный вывод: всякий генотип имеет хотя и большие, но вполне конкретные возможности мутационных изменений, определенный спектр изменчивости. Одни и те же мутации с определенной регулярностью появляются в пределах популяций данного вида вновь и вновь. И с другой стороны, некоторые изменения, кажущиеся столь же вероятными, никогда у этого вида не наблюдаются. Например, среди различных видов мух-Дрозофил, вероятно наиболее изученных в генетическом отношении животных, никогда не наблюдались особи с зелеными или синими глазами. Поскольку генетиками изучено уже около двух миллиардов (!) мух, вероятность обнаружения указанных мутаций в будущем выглядит ничтожной. По-видимому, в геноме дрозофил нет каких-то предпосылок, необходимых для возникновения таких мутаций, хотя подобные окраски глаз нередко наблюдаются у других видов насекомых. Для дрозофил эти мутации являются по каким-то причинам невозможными ("запрещенными").
Мутации могут происходить на разных уровнях организации наследственного аппарата: на уровне полного хромосомного набора (генома) клетки, на уровне его морфологически обособленных частей -- хромосом и на уровне его структурно-функциональных единиц - генов.
Геномные мутации представляют собой изменения количества хромосом, которые могут происходить либо путем кратного увеличения числа целых (гаплоидных) наборов хромосом, либо посредством изменения числа отдельных хромосом в геноме, в результате чего общее число хромосом становится не кратным гаплоидному.
Увеличение числа целых хромосомных наборов (плоидности) в клетке называется полиплоидизацией, изменение числа отдельных хромосом в геноме - анеуплоидией, или гетероплоидией. Фенотипический эффект геномных мутаций основан на взаимодействии генов или их комплексов, расположенных в соответственных (гомологичных) хромосомах, число которых изменилось в результате данной мутации. Вероятно, это взаимодействие осуществляется через посредство продуктов белкового синтеза (ферментов) или РНК, транскрибированной с ДНК соответствующих хромосом.
Геномные мутации обычно приводят к стерильности мутантов в результате невозможности осуществления полового процесса с представителями родительской формы, сохранившими нормальное количество хромосом. Поэтому у животных относительно редко геномные мутации дают начало новым формам - это возможно только при способности данного вида размножаться бесполым способом, или посредством партеногенеза, или гиногенеза (гиногенез - способ размножения, при котором сперматозоид проникает в яйцеклетку и стимулирует ее развитие, но его ядро не сливается с ядром яйца и не участвует в развитии зародыша). Среди растений бесполое размножение распространено гораздо шире, чем среди животных. В связи с этим у растений полиплоидизация, вероятно, играла существенную роль в эволюционном процессе.
Обычно различают автополиплоидию и аллополиплоидию. Автополиплоиды образуются посредством увеличения числа хромосомных наборов у отдельных особей одного вида. У автополиплоидов каждая хромосома представлена одним и тем же числом гомологов, например в клетках тетраплоидов имеется по четыре гомологичные хромосомы, у октоплоидов - по восемь и т. д. Ал-лополиплоиды образуются посредством гибридизации организмов разных видов. У аллополиплоидов соответствующие хромосомы уже в определенной степени утратили свою гомологичность. Поэтому процесс мейоза, в котором осуществляется редукция числа хромосомных наборов перед половым процессом, у аллополиплоидов обычно нарушен. Аллополиплоиды могут сохраниться как новый вид, если способны к бесполому размножению.
Половой процесс может восстановиться у полиплоидной формы, когда в популяции накапливается достаточное количество полиплоидных особей. У аллополиплоидов, кроме того, необходимо еще дополнительное кратное удвоение числа хромосом (посредством эндомитоза в соматических клетках или нерасхождения хромосом при образовании гамет). (Эндомитоз - удвоение числа хромосом внутри ядра без образования веретена деления клетки). Тогда в аллополиплоидном наборе появляются гомологичные хромосомы, и в дальнейшем мейоз может протекать нормально. Известно несколько примеров возникновения новых форм растений этим путем; среди них можно упомянуть рафанобрассику - межродовой гибрид, полученный Г.Д.Карпеченко путем скрещивания редьки (Raphanus sativus) с капустой (Brassica oleracea).
Хромосомные мутации являются структурными перестройками отдельных хромосом, изменяющими расположение в них генов, фенотипическое проявление подобных мутаций зависит от изменений пространственной локализации генов в хромосомах (эффект положения). Выделяют несколько основных типов хромосомных мутаций: а) транслокация - взаимный обмен участками между двумя разными (не гомологичными друг другу) хромосомами; б) делеция - утрата хромосомой какого-либо ее участка; в) инверсия - поворот внутри хромосомы какого-либо ее участка на 180°; г) дупликация - удвоение тех или иных участков хромосомы; д) транспозиция - перестановка генов внутри хромосомы.
Среди указанных типов хромосомных мутаций особую роль играют дупликации, которые (так же как и полиплоидизация) позволяют увеличить количество генетического материала в клетках. Как подчеркнул С. Оно, возникновение избыточности генетического материала может играть важную роль в эволюционных перестройках генетической системы вида, открывая новые возможности для прогрессивной эволюции, в частности для развития качественных новообразований. Дупликации ведут к появлению в наследственном аппарате генов-дублеров, отвечающих за одну и ту же функцию. В таких дублерах могут в скрытой форме сохраняться и накапливаться новые генные мутации. С другой стороны, инверсии способствуют образованию устойчивых генных комплексов (групп сцепления), которые редко разрушаются кроссинговером при мейозе и обеспечивают совместное наследование различных признаков.
Наконец, генные или точковые мутации представляют собой изменения самих генов, т.е. элементарных функциональных единиц аппарата наследственности. Сущность генных мутаций заключается в изменении порядка пуриновых и пиримидиновых оснований в молекуле ДНК. Для возникновения одной точковой мутации достаточно изменения хотя бы одной пары оснований. Если такие мутации происходят в структурных генах, может измениться порядок считывания наследственной информации при транскрипции ("мутация рамки" - смешение рамки считывания наследственной информации), что приведет к изменению порядка аминокислот в полипептидной цепи, т. е. к синтезу нового типа белковых молекул. Мутации генов-регуляторов изменяют соотношения между репрессированными и активными структурными генами, что неизбежно влечет за собой изменения белковых комплексов, синтезируемых в различных типах соматических клеток, несмотря на оставшиеся неизменными структурные гены.
Генные мутации являются основным источником наследственной изменчивости организмов. В результате мутирования возникают различные варианты генов - аллели, обусловливающие разные состояния контролируемого данным геном признака (или признаков). Число аллельных состояний отдельных генов может быть очень большим (более ста).
Поскольку мутационная изменчивость неопределенна по отношению к характеру изменений внешней среды, вызвавших появление мутаций, подавляющее большинство возникших наследственных изменений не улучшают, а ухудшают приспособленность мутантных организмов, и тем в большей мере, чем больше масштаб фенотипического проявления мутации. Поэтому в среднем наибольшие шансы на сохранение имеют мутации с наименьшим фенотипическим выражением ("малые мутации"), которые и служат основным материалом эволюционных изменений.
Фенотипическое проявление действия генов
В гаплоидном состоянии клетка обладает лишь одним полным хромосомным набором, и каждый ген представлен каким-то одним аллелем. В этом случае любая мутация сразу проявляется фенотипически и сразу же подвергается действию отбора. Гаплоидия как состояние, обеспечивающее возможность в первом же поколении опробовать новые мутации фенотипически, убыстряет эволюционный процесс и повышает скорость развития новых приспособлений, но требует огромных потерь мутантных особей в каждом поколении, так как подавляющее большинство мутантов оказываются хуже приспособленными к существующим условиям. Поэтому гаплоидное состояние как основная форма существования вида возможно только у низших одноклеточных организмов, для которых характерны гигантская численность особей и огромные темпы размножения (бактерии, синезеленые водоросли, простейшие и др.). У таких организмов выбраковка непригодных мутантов не угрожает сохранению вида как целого, и в каждом поколении новые мутации сразу испытываются отбором. Это и обусловливает поразительную быстроту приспособления подобных организмов к изменениям условий (например, приспособления многих болезнетворных микроорганизмов к действию новых лекарственных препаратов).
Однако выгоды гаплоидного состояния не могут быть использованы более сложными многоклеточными организмами, популяции которых состоят из меньшего числа особей (численность многих видов позвоночных животных измеряется лишь тысячами особей), имеют длительный онтогенез и низкие темпы размножения. Большая выбраковка особей в каждом поколении для таких форм была бы гибельной. Вместе с тем наследственная изменчивость и неизбежна у любого вида организмов, и необходима для его выживания в изменяющихся условиях. Макроорганизмы нашли оптимальное решение этой проблемы в диплоидном состоянии, при котором в каждой соматической клетке имеется по два полных хромосомных набора, состоящих из гомологичных хромосом, которые несут гомологичные гены, отвечающие за одни и те же признаки. Наличие в обеих гомологичных хромосомах одинаковых аллелей какого-либо гена называется гомозиготным состоянием (по данному гену), наличие же разных аллелей - гетерозиготным состоянием. При гетерозиготности фенотипически проявляется лишь один из аллелей - доминантный, или же в фенотипе наблюдается особое состояние признака, отличающееся от гомозиготных фенотипов обоих аллелей (неполное доминирование). Благодаря таким соотношениям разных аллелей в диплоидном генотипе в гетерозиготном состоянии могут сохраняться в скрытой, не проявляющейся фенотипически (рецессивной) форме любые новые аллели. Так, новые мутации могут постепенно накапливаться в популяции данного вида в форме рецессивных аллелей, время от времени проявляясь фенотипически в гомозиготном состоянии и подвергаясь в этом случае действию отбора.
Новые мутации могут, конечно, сразу обладать доминантным проявлением в фенотипе диплоидных особей, но, как указано выше, вероятность выживания мутантов вообще невелика, и поэтому преимущественно сохраняются именно рецессивные мутации. Впоследствии, если при каких-либо изменениях внешних условий новый признак окажется благоприятным, обусловливающий его мутантный аллель может вторично приобрести доминантное фенотипическое выражение (следует подчеркнуть, что Доминантны и рецессивны, собственно говоря, не сами аллели, а их проявления в фенотипе). Переход аллеля от рецессивного к Доминантному состоянию может быть обусловлен различными механизмами, действующими на разных уровнях преобразований наследственной информации в онтогенезе. Генетически такой переход может быть достигнут через отбор особых генов-модификаторов, влияющих на фенотипическое проявление мутантного аллеля (гипотеза Р.Фишера), или же через отбор аллелей с большей физиологической активностью (обеспечивающих более интенсивный синтез ферментов), чем первоначальный рецессивный вари ант (гипотезы С.Райта и Д.Холдейна). В сущности, эти гипотезы не исключают, а взаимно дополняют друг друга, и эволюция доминантности может происходить путем отбора малых мутаций как структурных генов, так и генов-модификаторов.
Так или иначе степень доминантности фенотипического проявления аллелей может эволюционировать, повышаясь под контролем отбора, если данный аллель становится благоприятным для его носителя при изменениях внешних условий. Примером этого может служить повышение доминантности аллеля, контролирующего темную окраску бабочек березовой пяденицы (Biston betu-laria), которое произошло в течение последних ста лет в индустриальных районах Европы (явление, получившее название "индустриальный меланизм"; подробнее см. в гл. 3).
Соотношение фенотипической изменчивости, входящей в норму реакции генотипа, с изменчивостью наследственной, вообще говоря, достаточно сложно. Это соотношение носит в известном смысле парадоксальный характер: наследуются изменения генотипа (мутации), но отбор не может непосредственно их контролировать - отбор имеет дело с фенотипами организмов, в каждом из которых наследственная основа осуществила лишь один из многих возможных вариантов своего выражения. Другими словами, генотипическая изменчивость проявляется и подвергается отбору только в форме фенотипических признаков. При этом важно подчеркнуть, что отбор, действуя на фенотипы организмов, способствует выживанию или же элиминирует целостные генотипы, обладающие определенной нормой реакции.
Взаимодействие различных генов в целостном генотипе существенно влияет на общую норму реакции генотипа. В зависимости от этого физиологическое проявление одного и того же гена в одних и тех же внешних условиях может быть различным у особей с разными генотипами, т.е. в разной генетической среде. Данный ген может проявляться как доминантный у одних особей и не проявляться фенотипически у других. Н.В.Тимофеев-Ресовский назвал это свойство пенетрантностью проявления гена. Пенетрантность мутантного гена измеряется процентным соотношением особей, имеющих мутантный фенотип, и общего количества особей-носителей данного аллеля в популяции. Если этот фенотип обнаруживается у всех носителей данного гена, говорят о полной (100%-ной) пенетрантности, если же лишь у некоторых особей - о неполной пенетрантности данного гена.
Пенетрантность генов зависит как от взаимодействия генов в генотипе, так и от внешних условий. При разных условиях у особей со сходным генотипом фенотипическое выражение данного гена может быть различно. Так, мутация дрозофилы "abnormal abdomen" (фенотипически проявляется в неправильном расположении полосок окраски на брюшке) обнаруживается в фенотипе только при выращивании личинок мух на влажном корме; при использовании сухого корма мутанты ничем не отличаются от нормальных мух.
Кроме того, может варьировать (даже при 100%-ной пенетрантности) степень фенотипического проявления гена. Признак может быть выражен сильнее или слабее, проявляться более или менее отчетливо в зависимости от внешних условий или от взаимодействия генов друг с другом. Изменчивость степени выраженности признака в фенотипе называют его экспрессивностью. Если данный признак сходно выражен у разных особей в данных условиях, говорят о постоянной экспрессивности контролирующего этот признак гена, если же степень выраженности признака варьирует - об изменчивой экспрессивности. Недавно было показано, что на экспрессию структурных генов могут оказывать влияние повторяющиеся нуклеотидные последовательности "молчащей ДНК", которая, таким образом, также воздействует на признаки фенотипа.
Р. Гольдшмидт обратил внимание на удивительный параллелизм между фенотипическими проявлениями модификационной и наследственной изменчивости. Одно и то же состояние фенотипических признаков у разных особей данного вида может сформироваться по-разному: у одних особей - как модификацион-ная вариация, проявляющаяся лишь в определенных условиях, у других - как единственно возможный вариант признака (т.е. для Данного аллеля со 100%-ной пенетрантностью и постоянной экспрессивностью). Если в популяции, где данный признак первоначально был одним из модификационных вариантов нормы реакции, появляется мутация, обусловливающая более высокую Пенетрантность и экспрессивность развития этого признака, возникает иллюзия "превращения" ненаследственной модификации в наследственный признак ("эффект Болдуина"), т. е. "унаследования приобретенного соматического признака" в духе ламаркизма. о Действительности здесь имеет место просто возникновение новой мутации (рис. 7) - либо структурного гена, либо гена-модификатора, влияющего на фенотипическое проявление данного признака.

Модификации, сходные с фенотипическим проявлением определенных мутаций, получили название фенокопии мутаций; соответствующие мутации, "дублирующие" модификации, называются генокопиями последних. Так, у плодовой мушки-дрозофилы мутация tetraptera, при которой вместо обычной пары крыльев развиваются две пары (рис. 8), является генокопией модификации-морфоза, возникающей при обработке ранних личиночных стадий эфиром или высокой температурой. Соответственно этот морфоз является фенокопией указанной мутации.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 611; Нарушение авторских прав?; Мы поможем в написании вашей работы!