КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Часть II. Микроэволюция и видообразование 3 страница
|
|
|
|
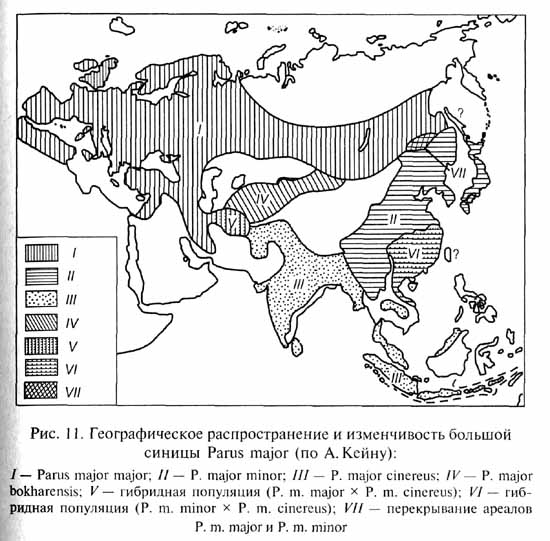
В некоторых случаях, особенно у видов с достаточно большим ареалом, может возникнуть еще более любопытная ситуация, которую хорошо иллюстрирует следующий пример. Большая синица (Parus major) широко распространена в Евразии - от Ирландии до Японии и Индонезии (рис. 11). В пределах этого огромного ареала орнитологи выделяют свыше 30 подвидов большой синицы. Все подвиды, соседствующие друг с другом, легко скрещиваются с образованием зон интерградации. Расселение большой синицы на восток осуществлялось, по-видимому, по двум направлениям: северному (леса умеренного пояса, к северу от зоны пустынь и гор Центральной Азии) и южному (Персия, тропические леса Индии и Индокитая). На Амуре встретились две формы большой синицы, проникшие сюда по северному и южному путям расселения: подвид Parus major major (распространенный от Западной Европы до Дальнего Востока) и подвид P. major minor. Эти две формы не скрещиваются друг с другом, т.е. ведут себя как самостоятельные виды. Однако P. m. minor легко скрещивается с более Южными подвидами большой синицы, а те, в свою очередь, - с подвидами, обитающими в Индии и Персии; последние формы легко скрещиваются с европейскими P. m. major. Как же приложить определение биологического вида к этой причудливой картине родственных форм? С одной стороны, P. m. major и Р. m. Minor по их нескрещиваемости друг с другом можно оценить как два разных вида; с другой стороны, они связаны через цепь легко скрещивающихся южных популяций, по которым в принципе возможна передача генетической информации от одного конца цепи до другого. Подобные ситуации встречаются не столь уж редко, как можно было бы подумать. Б. Ренш предложил обозначать такую цепь родственных аллопатрических форм термином "круг рас", а если некоторые формы в этом "круге" заслуживают выделения в качестве самостоятельных видов - "кольцевой вид". Позднее Э.Майр назвал такие аллопатрические родственные формы, которые на основании имеющихся данных трудно определить как подвиды или самостоятельные виды, "полувидами", а группу родственных полувидов (т.е. кольцевой вид Б.Ренша) - "надвидом".
Вид во времени
Вторая неопределенность в концепции биологического вида выявляется при попытке приложить определение Майра к популяциям, разделенным не географически, а во времени, т.е. к популяциям, связанным друг с другом родством не по горизонтали, а по вертикали, как предки с потомками. За истекший промежуток времени вид может существенно измениться. Так как изменения генофонда постепенны, невозможно провести отчетливую границу между предковым видом и любым из видов-потомков, если известен весь эволюционный ствол. Эта проблема неизбежно встает перед палеонтологами при изучении групп организмов, Достаточно полно представленных в палеонтологической летописи. Выход можно отчасти найти, используя ряд дополнительных терминов. Непрерывный ряд последовательных во времени популяций, каждая из которых является потомком предшествующей и предком последующей, называют филетической линией. Случайные отрезки этой последней, обычно известные палеонтологам, представляют собой палеонтологические виды (см. рис. 10). Наконец, биологический вид - это отдельная филетическая линия, рассматриваемая в данный момент времени, включая все живущие одновременно популяции, соответствующие определению Э. Майра. Биологический вид является, так сказать, "срезом" филетической линии плоскостью времени, или филетической линией, рассматриваемой в течение кванта времени (А.Кейн).
Д. Симпсон предложил для наименования отдельной филетической линии особый термин " эволюционный вид ", дополнив определение филетической линии указанием на ее эволюционную независимость от других линий, собственную эволюционную роль и определенную тенденцию эволюционных изменений. Эта концепция подчеркивает эволюционную целостность вида и может быть использована в анализе филетических линий, переплетающихся друг с другом или расходящихся как независимые ветви.
Приспособленность и приспособляемость вида
Как уже отмечалось, внутривидовая популяционная организация является приспособлением к оптимальному использованию всего разнообразия условий в пределах видового ареала. Приспособленность всякой популяции к конкретным условиям ее обитания определяется специфическим подбором аллелей в ее генофонде и в результате этого - соответствующей нормой реакции особей, обусловливающей определенные фенотипические особенности. Различия популяций выражаются прежде всего в характере поведенческих реакций особей, совокупность которых определяет такие важные экологические характеристики популяции, как особенности питания, плотность населения, возрастная и половая структура, сезонные реакции, динамика численности и др.
Действие отбора придает указанным особенностям популяций черты целесообразности. Благодаря отбору одновременно с приспособленностью к существующим условиям популяции приобретают также определенную степень приспособляемости, т.е. способность выдержать разного рода изменения условий. Приспособляемость популяций определяется, с одной стороны, широтой нормы реакции составляющих популяцию генотипов (т.е. возможностями их модификационной изменчивости), с другой же стороны - генетической гетерогенностью, разнообразием аллелей в пределах популяционного генофонда (т.е. запасом комбинативной изменчивости). Важность этого мобилизационного резерва изменчивости для выживания популяций и эволюционную необходимость высокой степени генетической гетерогенности природных популяций показал С.С.Четвериков. При изменениях условий, выходящих за рамки нормы реакции генотипов, которые были оптимально приспособлены к прежним условиям, популяция сможет сохраниться только при наличии в ее генофонде таких генотипов, которые окажутся жизнеспособными и в новых условиях. Следовательно, отбор благоприятствует поддержанию генетической гетерогенности популяций. Запас комбинативной изменчивости включает наряду с оптимальными в данных условиях аллелями также относительно слабо приспособленные аллели. Последние снижают общую приспособленность популяции и составляют так называемый генетический груз, который, по образному выражению Д.Холдейна, является той ценой, которую вынуждена платить популяция за право эволюционировать и, добавим, чтобы выжить и сохраниться во времени. Благодаря этому приспособляемость популяции, которой благоприятствует генетическая гетерогенность и, как следствие, генетический груз, до известной степени противостоит приспособленности популяции к существующим условиям, основанной на оптимальных аллелях. Таким образом, в природных популяциях должно существовать некоторое равновесие между этими двумя противоположными тенденциями.
По определению Т. Мёллера, генетический груз представляет собой ту часть популяционного генофонда, которая определяет появление менее приспособленных особей, подвергающихся избирательной выбраковке (элиминации) под воздействием естественного отбора. Общий генетический груз складывается из нескольких составляющих: 1) мутационный груз -- неудачные мутации, существенно снижающие приспособленность; 2) сегрегационный груз - выщепляющиеся в каждом поколении слабо приспособленные гомозиготные особи по таким аллелям, которые в гетерозиготном состоянии обладают высокой приспособленностью (так называемое "сверхдоминирование", см. ниже); 3) субституционный груз - результат изменений приспособленности фенотипи-Ческих признаков при изменениях внешних условий или появлении новых, более удачных фенотипических вариантов.
Эволюционные преобразования вида под воздействием отбора Неизбежно сопровождаются гибелью хуже приспособленных особей. Общее число отбракованных отбором особей при замещении одного аллеля другим Д.Холдейн назвал "платой за отбор". По Расчетам Холдейна, плата за отбор при одновременном эволюционном замещении нескольких независимых аллелей (разных генов) оказывается чрезвычайно высокой: она экспоненциально возрастает с увеличением числа независимых генов, подвергающихся отбору. Из этого был сделан парадоксальный вывод о невозможности быстрых эволюционных преобразований, основанных на одновременном замещении многих независимых аллелей - которые, однако, реально происходили в филогенезе многих групп организмов. Этот парадокс решается, если учесть, что отбор действует, как уже подчеркивалось, не на отдельные гены, а на фенотипы организмов, формирующиеся в онтогенезе на основе взаимодействий целостных генетических систем ("супергенов") в целостном генотипе. В результате отбору подвергаются не отдельные гены, а полигенные комплексы (супергены), и эволюционные преобразования затрагивают сопряженные комплексы фенотипических признаков. Это и определяет возможность высокой скорости эволюции без гибельных последствий для вида в результате слишком высокой платы за отбор (подробнее см. кн.: Грант В. Эволюция организмов. - М, 1980).
Полиморфизм вида
Существующие в данный исторический момент условия среды могут благоприятствовать либо какому-то одному фенотипу, либо сразу нескольким разным фенотипам. В первом случае в природных популяциях преобладает один доминантный фенотип, называемый "диким типом", который скрывает в гетерозиготном состоянии многочисленные рецессивные аллели (таков, например, хорошо известный фенотип окраски домовой мыши). Во втором же случае в одной популяции постоянно встречаются несколько устойчивых, отчетливо различающихся друг от друга форм. Такое состояние называется полиморфизмом популяции (вида).
Очевидно, полиморфизм позволяет популяции лучше использовать разнообразие условий, существующих в пределах ее ареала аналогично тому, как дифференциация аллопатрических популяций позволяет виду лучше использовать разнообразие условий в пределах видового ареала. Полиморфизм повышает одновременно и приспособленность, и приспособляемость популяции, поскольку он всегда основывается на генетической гетерогенности, на существовании в популяции нескольких устойчивых генных комплексов.
Полиморфизм может и не иметь отчетливого приспособительного характера, основываясь на адаптивной равноценности рач-ных вариантов (см. в гл. 1 о полиморфизме белковых комплексов). Но здесь нас интересует в первую очередь адаптивный, сбалансированный отбором полиморфизм. Такой полиморфизм в популяции возникает в следующих основных случаях: 1) отбор действует против фенотипа, обусловленного гомозиготным состоянием определенного аллеля, но благоприятствует гетерозиготному фенотипу, обладающему повышенной жизнеспособностью ("сверхдоминирование" - гетерозиготы превосходят по жизнеспособности оба вида гомозигот); 2) на различных стадиях онтогенеза отбор благоприятствует фенотипическим особенностям, обусловленным разными аллелями; 3) отбор благоприятствует сохранению данного аллеля у особей одного пола, но действует против него у особей другого пола; 4) в разные сезоны года более приспособленными оказываются фенотипы, обусловленные разными аллелями; 5) фенотипы, обусловленные разными аллелями, используют различные условия в пределах одного ареала; 6) отбор благоприятствует сохранению данного фенотипа, когда этот последний редок, и направлен против него, когда он встречается часто.
Явления возрастного полиморфизма и полового диморфизма (вторая и третья формы полиморфизма в этом перечне) хорошо известны. Остальные варианты рассмотрим подробнее. Интересным примером "сверхдоминирования" является полиморфизм некоторых восточноафриканских популяций человека по типам гемоглобина. В этих популяциях у взрослых людей кроме обычного типа гемоглобина А наблюдается также особая форма гемоглобина S, которая отличается от типа А тем, что в одной из его молекулярных цепей место глютаминовой аминокислоты занимает валин. Нормальный аллель НbА встречается с частотой около 80%, мутантный аллель Hbs - около 20%. У гетерозигот НbА Hbs имеются обе формы гемоглобина, а у гомозигот HbsHbs почти весь гемоглобин крови представлен формой S. В этом последнем случае у людей развивается тяжелое заболевание - серповидно-клеточная анемия (эритроциты сморщиваются, принимая форму серпа), часто заканчивающаяся смертью. При таком исходе следовало бы ожидать гораздо меньшей частоты встречаемости мутантного аллеля, чем реально наблюдаемые 20%. Оказалось, что полиморфизм популяций по указанным аллелям связан с повышенной устойчивостью гетерозигот HbAHbs к малярии: количество паразитов в крови гетерозиготных индивидов много ниже, и продолжительность болезни короче, чем у гомозигот НbAНbА. Поскольку малярия является опасным заболеванием, вызывающим значительную смертность среди детей в Восточной Африке, очевидно, что распространение мутантного аллеля Hbs в человеческих популяциях в определенной концентрации будет выгодно, несмотря на опасность смертности гомозигот HbsHbs от анемии. В результате аллель Hbs имеет повышенную концентрацию именно в тех районах, где наиболее распространена малярия.
Наземные улитки Cepaea nemoralis в зоне смешанных лесов Умеренного пояса характеризуются сбалансированным полиморфизмом окраски раковины: выделяют бурую, розовую и желтую формы. Ранней весной наименее заметными на преобладающем фоне окружающей среды оказываются раковины бурой и розовой окраски, летом - желтые раковины. В то же время желтая форма менее заметна среди луговой растительности, тогда как бурая и розовая - в лесу. Очевидно, полиморфизм окраски у улиток соответствует 4-й и 5-й формам полиморфизма в приведенном выше перечне.
Наконец, последняя из указанных форм полиморфизма наблюдается, например, при мимикрии (подражательном сходстве) какого-либо вида с несъедобным для хищников видом-моделью. Такое сходство благоприятно для вида-подражателя, только если численность мимикрирующего фенотипа в его популяциях будет существенно ниже численности несъедобного вида-модели (в противном случае хищник не сможет эффективно "обучаться" не трогать особей с данным фенотипом). В ситуации, представленной на рис. 5, среди самок бабочки Papilio dardanus, разные цветовые устойчивые формы (морфы) имитируют окраску разных несъедобных бабочек-данаид, тогда как самцы P.dardanus вообще не мимикрируют.
Симпатрические внутривидовые группировки
Полиморфизм популяций является основой для возникновения в некоторых случаях устойчивых и обособленных внутривидовых группировок особей ("биологических популяций"), обитающих в одном и том же районе, т.е. обладающих симпатрическим распространением, в отличие от аллопатрических территориальных популяций. К ним относятся, в частности, возрастные и половые группы некоторых видов. Биологические популяции обособляются друг от друга, обитая в пределах одного ареала, либо в результате специфических взаимоотношений с внешней средой, либо благодаря особенностям внутривидовой организации. Примеры подобной внутривидовой дифференциации представляют обособленные друг от друга биологически личиночные и взрослые популяции земноводных или насекомых с полным превращением, или же такие связанные с особенностями размножения группировки, как "гаремы" и стада самцов-холостяков, характерные для целого ряда видов млекопитающих (кашалоты, олени, зебры, куланы и др.).
Особой формой биологических популяций являются так называемые биологические расы. Этим термином обозначают такие устойчивые группировки особей одного вида, которые имеют симпатрическое распространение, но различаются друг от друга определенными экологическими особенностями (например, характером питания, сроками и местами размножения и т.п.). Благодаря этим различиям разные биологические расы одного вида держатся раздельно друг от друга, хотя и имеют общий ареал. В результате обмен генетической информацией между разными биологическими расами одного вида может быть снижен, как и между разными территориальными (аллопатрическими) популяциями.
Примером подобных биологических рас могут служить так называемые сезонные ("яровые" и "озимые") расы ряда видов лососевых и осетровых рыб, описанные Л. С. Бергом. Речь идет о проходных рыбах, которые обитают в море, но для икрометания заходят в реки. Сезонные расы описаны Бергом у кеты (Oncorhyn-chus keta), чавычи (О. tschawytscha), нерки (О. nerka), семги (Salmo salar), каспийского лосося (S.trutta caspius), севрюги (Acipenser stellatus) и других видов. Рыбы "озимой" расы входят в реки осенью, зимой держатся в их низовьях, а рано весной поднимаются вверх по течению для нереста. Рыбы "яровой" расы проводят зиму в море и входят в реки позднее - когда нерест рыб "озимой" расы уже заканчивается. Благодаря этому обе расы одного вида, обитая в одном географическом районе и выметывая икру в одних и тех же реках, в значительной степени изолированы друг от друга во время сезона размножения и обмен генетической информацией между ними затруднен.
Другой пример представляют биологические расы обыкновенной кукушки (Cuculus canorus), описанные А.С.Мальчевским. Как известно, кукушка откладывает яйца в гнезда различных видов воробьиных птиц (это явление иногда называют "гнездовым паразитизмом"). Представители разных биологических рас обыкновенной кукушки проявляют предпочтение к "своему" виду воробьиных, в гнезда которого они откладывают яйца. При этом окраска яиц у кукушек разных рас соответствует окраске яиц того вида птиц, на котором "паразитирует" данная раса кукушек. Следовательно, в этом случае разные биологические расы проявляют уже Достаточно отчетливые наследственные различия морфологического признака (окраска яиц).
Очевидно, биологические внутривидовые группировки, так же как и территориальные популяции, позволяют виду более полно и совершенно использовать ресурсы в пределах занимаемого им ареала.
Подводя итоги, мы можем с полным основанием констатировать, что биологический вид представляет собой сложную систему, обладающую высокой устойчивостью к изменениям внешней среды и приспособленную к оптимальному использованию существующего разнообразия условий. Высокая степень приспособленности и приспособляемости вида обеспечивается несколькими путями, включающими мутационную, комбинативную и моди-фикационную изменчивость и формирующиеся на базе комбинативной изменчивости политипичность и полиморфизм. Вид представляет собой систему с нежесткими связями взаимозаменяемых компонентов (так сказать, с "параллельным включением" звеньев), лишенную какого-либо интегрирующего, контролирующего всю систему центра. Эта сложная динамическая система возникает, поддерживается и совершенствуется под контролем естественного отбора.
ГЛАВА 3. ФАКТОРЫ ЭВОЛЮЦИИ
В предыдущих главах мы рассмотрели материальные предпосылки эволюции (наследственную изменчивость) и биологические макросистемы - виды и популяции, являющиеся структурными единицами эволюционного процесса. Теперь мы можем перейти к рассмотрению движущих сил эволюции, важнейшей из которых является естественный отбор.
Естественный отбор
Как уже было отмечено, мутационная изменчивость необходима, но сама по себе еще недостаточна для осуществления приспособительной и прогрессивной эволюции. Приспособленность к конкретным условиям существования - важнейшая предпосылка выживания и нормальной жизнедеятельности организмов - формируется только благодаря действию естественного отбора. Естественный отбор является творческой силой эволюции, направляющей (векторизующей) эволюционный процесс и интегрирующей отдельные изменения в адаптации. В этом современные представления вполне соответствуют взглядам Дарвина. Однако следует отметить, что отбор не требует перенаселения как обязательного условия, обеспечивающего его эффективность. Борьба за существование, понимаемая в том широком и метафорическом смысле, который вкладывал в него сам Дарвин, означает всю совокупность взаимоотношений организма и внешней среды и, следовательно, должна иметь место и без перенаселения. Естественный отбор поэтому должен действовать и в ситуациях, далеких от перенаселения.
Генетическая сущность отбора заключается в неслучайном (избирательном) выживании изменчивых генотипов и в избирательном участии разных генотипов в передаче генов последующим поколениям. Следовательно, естественный отбор имеет два существенно различающихся аспекта: отбор на выживание, определяющий дифференцированное выживание разных особей, и репродуктивный отбор (не путать с половым отбором), обеспечивающий различную степень участия разных особей в размножении. Эти два аспекта отбора могут не совпадать - например, стерильный гибрид может обладать повышенной жизнеспособностью.
Различные факторы внешней среды, воздействующие на данный вид организмов и требующие определенной приспособительной реакции организмов, представляют собой отдельные силы отбора (или селективные силы). Общее воздействие отбора на данный вид организмов, иногда называемое давлением отбора или селективным давлением, складывается как результат интеграции отдельных селективных сил. Взаимодействие последних друг с другом может оказывать существенное влияние на общий эффект отбора.
Если разные селективные силы имеют несовпадающие направления, общий итог их взаимодействия получает характер эволюционного компромисса, подразумевающего формирование под контролем отбора такого фенотипа, который обеспечивает скорее некоторое оптимальное соотношение разных адаптации, чем достижение максимального совершенства одной из них.
Необходимо отметить, что до сих пор определенное распространение (особенно среди критиков дарвинизма) имеют упрощенные и, по существу, неверные представления о естественном отборе, которые ведут к серьезным ошибкам в понимании механизмов эволюционного процесса. Прежде всего неверно представление о естественном отборе как изолированном действии отдельных селективных сил на отдельные признаки организма, т. е. как отборе отдельных признаков (и тем более - отдельных аллелей). Темпы эволюции, рассчитанные на основе этих представлений, оказываются гораздо более низкими, чем наблюдаемые реально поданным палеонтологии темпы эволюции многих филетических линий, а плата за отбор - непомерно высокой. Напомним, что отбору непосредственно подвергаются не отдельные гены, а определенные фенотипы, формирующиеся в онтогенезе на основе соответствующих генотипов при модифицирующем влиянии внешних условий. Следовательно, отбираются целые генотипы, характеризующиеся такой нормой реакции, которая наиболее благоприятна в данных условиях среды по всей совокупности признаков. Благодаря блочно-иерархическому принципу организации генома отбор может действовать на целостные генетические комплексы (супергены), что на несколько порядков ускоряет эволюционный процесс.
Поскольку всякий генотип характеризуется специфическим спектром мутационных изменений (т. е. обладает способностью с некоторой частотой давать определенные мутации), естественный отбор, благоприятствуя конкретным генотипам, тем самым благоприятствует и соответствующему направлению и частоте возникновения наследственных вариаций. Таким образом, если отбор благоприятствует каким-то фенотипическим признакам, носителем которых является данный генотип, то отбор тем самым влияет и на характер дальнейших эволюционных изменений.
Преобладающие направления изменчивости различных генотипов могут в разной степени соответствовать направлению отбора. В этом случае при наличии двух генотипов, которые являются носителями какого-то нового полезного признака, но различаются своими спектрами изменчивости, отбор, очевидно, будет в большей мере благоприятствовать тому генотипу, возможности изменчивости которого будут ближе к направлению действия отбора. При совпадении преобладающего направления изменчивости и направления отбора приспособительная эволюция пойдет гораздо быстрее, чем при отсутствии такого совпадения. Быстрота возникновения и усовершенствования приспособлений дает очевидное преимущество в борьбе за существование. Таким же образом отбор может воздействовать и на частоту определенных мутаций (через посредство соответствующих генов-мутаторов), поскольку этот показатель существенно влияет на распространенность в популяции благоприятных вариантов.
Следовательно, естественный отбор, выбраковывая одни и сохраняя другие генотипы, тем самым оказывает влияние на направление и частоту возникающих наследственных изменений, что также способствует значительному ускорению эволюционного процесса.
Вторая группа ошибочных представлений о естественном отборе связана с непониманием его творческой роли. Эти представления коренятся в упрощенной концепции отбора, которая на первый взгляд кажется очевидной: отбор понимается как своего рода "сито", устраняющее наименее приспособленные варианты наследственных изменений (мутаций) и сохраняющее более приспособленные. Именно такое понимание отбора характерно, например, для сторонников "нейтральной эволюции" (М. Кимура, С. Оно и др.), подчеркивающих "консервативную природу" отбора и видящих движущую силу эволюции в мутационном процессе (см. гл. 1). Этот очевидный результат действия отбора, несомненно, существует, но отнюдь не исчерпывает его эффект во всей полноте. При концентрации внимания на "консервативной роли" отбора странным образом забывают, что отбор не только выбраковывает неудачные варианты, но и накапливает удачные, формируя из них адаптации любой степени сложности, и преобразует эти адаптации в соответствии с изменениями внешних условий.
Отбор оказывает на организм многостороннее интегрирующее влияние, способствуя оптимизации взаимодействий разных его систем. Приспособительное, формообразовательное и интегрирующее воздействие отбора на организм и обозначается как творческая роль естественного отбора.
Интенсивность действия отбора
Естественный отбор можно представить как вектор, поскольку его, с одной стороны, характеризует величина, которую можно выразить количественно, с другой стороны - направление.
Количественная характеристика действия отбора носит название его интенсивности или эффективности: она представляет собой степень воздействия (давления) отбора на популяцию. Очевидно, давление отбора в разных условиях может быть большим или меньшим. Для количественного выражения интенсивности отбора используют показатель, называемый коэффициентом отбора.
Пусть некоторая популяция включает два типа особей - А и В.
Концентрации особей А и В в популяции соотносятся как СА/СВ= u1.
Если один из этих вариантов имеет преимущество перед другим, измеряемое коэффициентом интенсивности отбора S, то в следующем поколении соотношение концентраций особей А и В в популяции изменится в соответствии с этим коэффициентом:
u2= u1(1-S)
S=(u1-u2)/u1=1-u2/u1
где u2 – соотношение концентрации особей в следующем поколении.
Если u1 > u2, то S >0 и особи В имеют какое-то преимущество перед особями А, которое "отмечается" отбором, и концентрация особей В в популяции возрастает от поколения к поколению. Если же u1=u2, то S = 0, следовательно, особи А и В равноценны и соотношение их концентраций в данной популяции не меняется.
Коэффициент интенсивности отбора позволяет сравнивать давление отбора на разные фенотипы. Для полиморфной популяции устанавливают коэффициент отбора всех вариантов по отношению к наиболее приспособленному (т.е. наиболее жизнеспособному и плодовитому в данных условиях) фенотипу. Очевидно, коэффициент отбора представляет собой величину относительную и не может характеризовать действие отбора только на данный фенотип без сравнения с другими вариантами.
Формы отбора
Если интенсивность давления отбора является его количественной характеристикой, то направление естественного отбора определяет качественную специфику его воздействия на популяцию. Выделяют три основных направления действия отбора, о которых обычно говорят как о типах (или формах) естественного отбора.
Стабилизирующий отбор. Если условия внешней среды длительное время остаются стабильными, приспособленность вида к этим условиям постепенно совершенствуется, пока не достигнет определенного оптимума. Совершенство всякого приспособления, разумеется, относительно, обычно возможна и более высокая степень приспособленности к данным условиям. Однако эволюционные возможности всякого конкретного генофонда, приспосабливаемого отбором к данным условиям среды хотя и велики, но ограничены спектром возможных мутационных изменений. Поэтому раньше или позже при постоянстве условий среды и при отсутствии конкуренции с экологически близкими формами генофонд данного вида оптимизируется по отношению к существующим условиям. С этого момента на популяцию начинает действовать стабилизирующий отбор, благоприятствующий сохранению оптимального генофонда, на базе которого формируются оптимально приспособленные к данным условиям фенотипы, которые становятся преобладающими в популяции. Процессы онтогенетического формообразования (морфогенезы), основывающиеся на разных генотипах, направляются стабилизирующим отбором в определенные русла, ведущие к формированию оптимального фенотипа (посредством распространения в популяции соответствующих генов-модификаторов). К.Уоддингтон назвал такое состояние канализованностью процессов морфогенеза.
В результате дикий тип, т.е. преобладающий в природной популяции оптимальный фенотип (или несколько фенотипов в полиморфной популяции), обычно объединяет генетически разнородных особей. Запас комбинативной изменчивости популяции благодаря канализованности процессов морфогенеза оказывается как бы скрытым за фенотипическим однообразием особей. Такое соотношение комбинативной и модификационной изменчивости оптимально при стабильных условиях среды, поскольку запас комбинативной изменчивости необходим для вида как "запас прочности", повышающий приспособляемость к изменениям среды обитания, а подгонка разных генотипов к одному или немногим оптимальным фенотипам повышает приспособленность к существующим в данный момент условиям. Состояние оптимальной стабильности популяции, которое обеспечивается ее специфическим генофондом и канализованностью процессов морфогенеза под контролем стабилизирующего отбора, носит название популяционного гомеостаза.
Стабилизирующий отбор направлен против крайних вариантов изменчивости, наблюдаемых в популяции, и благоприятствует консервативной части последней (рис. 12). В результате средняя величина (среднее состояние) признака остается из поколения в поколение неизменной. Стабилизирующий отбор ведет к большей фенотипической однородности популяции и к стабильности спектра ее изменчивости.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 909; Нарушение авторских прав?; Мы поможем в написании вашей работы!