КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Часть II. Микроэволюция и видообразование 6 страница
|
|
|
|
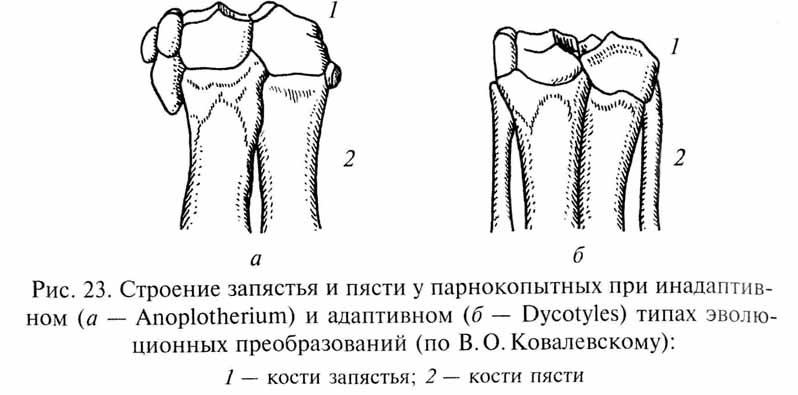
При анализе подобных ситуаций неизбежно возникает вопрос: как объяснить развитие инадаптивных изменений? Не противоречит ли "инадаптивная эволюция" представлениям о развитии приспособлений под контролем естественного отбора? Однако противоречия здесь нет. Дело в том, что на первых этапах эволюционных преобразований многих органов (в данном случае - в процессе редукции боковых лучей конечностей у парнокопытных) и адаптивные, и инадаптивные варианты изменений могут быть примерно в равной степени полезны для вида. Естественный отбор благоприятствует любому изменению, повышающему приспособленность организмов, вне зависимости от того, как пойдет развитие этого признака в дальнейшем. Строго говоря, термин В.О.Ковалевского "инадаптивная", т.е. неприспособительная, эволюция неудачен, так как отбираются только приспособительные (адаптивные) изменения организмов. Однако в ходе дальнейшей эволюции обнаруживается преимущество одного пути эволюционных преобразований перед другими. Механическое несовершенство конечностей "инадаптивных" форм копытных выявилось лишь при достаточно далеко зашедшей редукции боковых пальцев, а в начале этого процесса указанные формы, вероятно, ничем не уступали представителям "адаптивных линий".
Таким образом, можно назвать инадаптивным такой путь филогенетических изменений, при котором формирующееся приспособление заключает в себе предпосылки для возникновения тех или иных внутренних противоречий, которые нарастают мере развития приспособления, приводя в конечном итоге к эволюционному тупику. Как показал В.О.Ковалевский, на начальных стадиях развития инадаптивные изменения могут даже и определенные преимущества перед более медленно протекающими гармоническими преобразованиями - именно благодаря быстроте развития инадаптивных вариантов. Гармонические преобразования требуют большего числа эволюционных "проб и ошибок" и соответственно большего времени для своего осуществления. Вероятно, можно сказать, что быстрота эволюционных преобразований нередко оказывается обратно пропорциональной степени их совершенства.
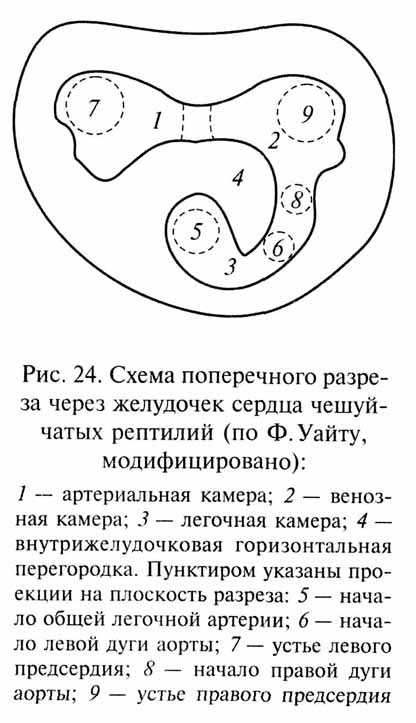
Инадаптивный путь эволюции не обязательно ведет к вымиранию - возникшие противоречия могут быть различными способами компенсированы. Рассмотрим, например, эволюционные преобразования сердца у рептилий завропсидного ствола (черепахи, клювоголовые, ящерицы, змеи, крокодилы). У предков завропсидных пресмыкающихся в желудочке сердца возникла горизонтальная перегородка, разделившая его на спинную и брюшную камеры и способствовавшая распределению артериальной крови в дуги аорты, а венозной - в легочную артерию (рис. 24). Однако полное разделение желудочка этой перегородкой, которое совершенно прекратило бы смешивание артериальной и венозной крови, невозможно - при этом легочная артерия, начинающаяся от брюшной камеры желудочка, оказалась бы отрезанной от обоих предсердий, открывающихся в спинную камеру. В результате у большинства пресмыкающихся перегородка в желудочке сердца остается неполной и сохраняется смешивание артериальной и венозной крови в сердце, что препятствует интенсификации метаболизма до уровня, необходимого для достижения гомойотермии (независимости температуры тела от температуры внешней среды). У предков крокодилов и птиц первичная горизонтальная перегородка в желудочке сердца редуцировалась и вместо нее возникла новая - вертикальная перегородка, полностью разделившая желудочек на левую (артериальную) и правую венозную) камеры. После редукции у предков птиц левой дуги начинавшейся от правой камеры желудочка, полностью прекратилось смешивание артериальной и венозной крови, что стало одной из важных предпосылок для развития у птиц гомойотермии. У предков млекопитающих в желудочке сердца сразу возникла вертикальная перегородка, которая, пока она была неполной, вероятно, не давала этим животным каких-либо преимуществ в разделении артериальной и венозной крови по сравнению с "типичными" пресмыкающимися - но этот путь эволюционных преобразований сердца позволил в дальнейшем разделить желудочек полностью без сложных дополнительных перестроек. Можно полагать, что предки завропсидных рептилий пошли по "инадаптивному" пути в разделении желудочка сердца по сравнению с более удачным ("адаптивным") решением этой проблемы у терапсидных рептилий - предков млекопитающих.
Инадаптивные изменения неизбежно должны быть обычным явлением в филогенезе различных групп организмов, ибо единственный фактор эволюции, обусловливающий развитие приспособлений, - естественный отбор - действует, так сказать, "в настоящем времени" и не может предвидеть эволюционных последствий преобразований, благоприятных в данный исторический момент и при данных условиях, но могущих при их дальнейшем развитии привести к эволюционному тупику и вымиранию.
Гиперадаптивность
На фоне обшей относительности совершенства приспособлений, наглядно проявляющейся в явлениях инадаптивной эволюции, особенно контрастно выглядит так называемая гиперадаптивность или конструктивная избыточность многих биологических систем, представляющая собой более или менее значительное (в некоторых случаях многократное) превышение их адаптивных возможностей по отношению к обычным условиям их функционирования. Это относится, например, к прочности костей на разрыв и сжатие (у млекопитающих она близка к прочности стали), к силе сокращения мышц (для одной мышцы лягушки она может примерно в 5 раз превышать вес всего тела) и т.п. Поразительную гиперадаптацию к действию низких температур проявляют в состоянии анабиоза споры ряда бактерий, а также некоторые микроскопические многоклеточные животные - коловратки (Rotatoria - класс типа нитчатых червей) и тихоходки (Tardigrada -- группа, близкая к кольчатым червям и членистоногим), которые могут выдержать температуру жидкого гелия (-270°С), близкую к абсолютному нулю.
Проявлением гиперадаптивности является также наличие в составе некоторых органов такого количества структурных элементов, которое во много раз превышает необходимое для их нормального функционирования при обычных условиях. Например, в женском яичнике закладывается от 40 до 200 тыс. яйцевых фолликулов, из которых овулирует в течение всей жизни женщины не более 500.
Важность и необходимость гиперадаптивности для выживания вида в изменчивой среде очевидна. В истории любого вида организмов неизбежно встречаются периоды необычных (экстремальных) условий, ставящих под угрозу существование целых популяций. Проявления гиперадаптивности представляют собой "запас прочности" систем организма, позволяющий им выдерживать повышенные функциональные нагрузки в таких экстремальных условиях. Конструктивная избыточность многократно повышает надежность биологических систем и является необходимым условием для выживания организмов, когда давление отбора достигает предельных значений.
Однако механизм возникновения гиперадаптивности является одной из сложных проблем адаптациогенеза. Естественный отбор, действуя лишь "в настоящем времени", может благоприятствовать только таким изменениям организмов, которые обеспечивают их носителям при данных условиях какие-либо преимущества в выживании или оставлении потомства. В то же время подавляющее большинство особей данного вида в течение всей жизни не подвергается воздействию экстремальных условий, при которых обладатели гиперадаптивного состояния получили бы соответствующие преимущества. Следовательно, гиперадаптивные признаки сами по себе имеют чрезвычайно мало шансов быть "замеченными" естественным отбором, и еще менее вероятны накопление и интеграция соответствующих изменений в ряду последовательных поколений. И совсем уже невероятным выглядит прямое закрепление отбором таких гиперадаптивных особенностей, которые обеспечивают сверхустойчивость организма (вплоть до условий, вообще не наблюдающихся ныне в биосфере Земли, например температур, близких к абсолютному нулю).
Чтобы разобраться в этой проблеме, прежде всего отметим, что понятие гиперадаптивности объединяет широкий круг очень разнородных явлений (в этом легко убедиться на приведенных выше примерах). Вероятно, возникновение разных форм конструктивной избыточности определяется различными причинами.
Вообще для нормального функционирования любого органа При самых обычных (средних) условиях полезно, чтобы диапазон его функциональных возможностей был существенно шире обычно используемого. Это позволяет избежать сбоев, вероятных в крайних областях функционального диапазона: краевые зоны неустойчивого функционирования отодвигаются за пределы обычно используемой "рабочей части" этого диапазона. Другими словами, некоторая потенциальная гиперадаптивность полезна уже для нормального функционирования органа в обычных условиях. Поэтому естественный отбор в процессе совершенствования любой адаптации благоприятствует расширению функциональных возможностей органа в гиперадаптивную область.
Дальнейшему усилению гиперадаптивности способствует действие катастрофического отбора (см. с. 108) в экстремальных условиях, раньше или позже неизбежно возникающих в истории любой филетической линии. При действии катастрофического отбора вид может выжить только в том случае, если в его популяциях имеются особи, обладающие соответствующей конструктивной избыточностью по сравнению с "нормой". При крайне неблагоприятных условиях катастрофический отбор элиминирует практически всю прежнюю популяцию, за исключением особей-носителей гиперадаптивного состояния отбираемых признаков. После возврата условий к прежней норме выжившие особи формируют новую популяцию, потомки которой будут сохранять конструктивную избыточность просто как своего рода нейтральные признаки (поскольку в нормальных условиях высокая степень гиперадаптивности отбором не контролируется) до следующей экстремальной ситуации. Очевидно, в длительной истории филетической линии важность гиперадаптаций для выживания вида не вызывает сомнений. Конструктивная избыточность, обеспечивающая запас прочности систем организма и дающая виду шансы на выживание в экстремальных условиях, является результатом действия катастрофического отбора в прошлой истории вида.
Кроме того, как показал П.В.Пучков, многие проявления конструктивной избыточности возникают как побочный результат контролируемого отбором развития обычных адаптации организмов к специфическим условиям их обитания. Так, гиперадаптивная устойчивость яиц жаброногих раков к действию низких температур (до -180°С), вероятно, возникла как побочный результат развития приспособлений, позволяющих этим организмам выживать при длительном пересыхании временных водоемов (благодаря особому химизму яиц и структуре их оболочек яйца жаброногих раков выдерживают высыхание до 9 лет), а также проходить без повреждений сквозь кишечный тракт проглотившего яйцо хищника.
Этот путь развития гиперадаптивных особенностей, очевидно, соответствует рассмотренному выше механизму морфофункиио-нальной преадаптации, обеспечивающему возможность расширения функций органов и приобретения ими новой адаптивной роли. Механизм преадаптации позволяет объяснить наиболее значительные проявления конструктивной избыточности организмов.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 658; Нарушение авторских прав?; Мы поможем в написании вашей работы!