КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Часть III. Историческое развитие (филогенез) организмов 2 страница
|
|
|
|
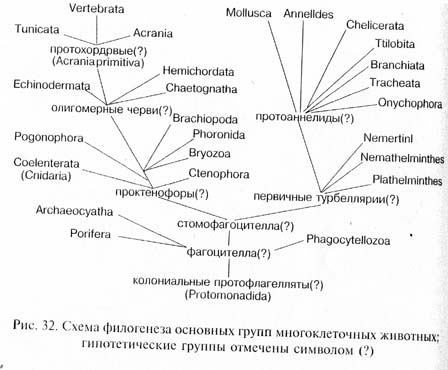
При дальнейшем развитии приспособлений к активному питанию происходила постепенная эпителизация фагоцитобласта, т.е. преобразование рыхлого скопления клеток в организованный клеточный пласт - эпителий кишечника. Эпителизация фагоиито-бласта, вероятно, началась с развития на заднем по движению полюсе тела постоянного ротового отверстия. Как отметил К. В. Беклемишев, на этой стадии филогенеза организм стал питаться как целое, а не как совокупность отдельных самостоятельно фагоцитирующих клеток. Вероятно, к этому времени сформировалась и интегрирующая организм нервная система в виде эпителиального нервного сплетения. Активное плавание требовало способности ориентироваться в пространстве и координировать работу всех органов. Для осуществления этих функций на аборальном (противоположном ротовому отверстию) полюсе тела возник нейро-рецепторный комплекс, включавший нервный ганглий, осязательные щетинки и статоцист (орган равновесия). Подобный аборальный орган имеется у современных гребневиков (тип Ctenophora), а также у свободноплавающих личинок очень многих групп животных: плоских и кольчатых червей, моллюсков, членистоногих, полухордовых, иглокожих и др. Эту гипотетическую стадию филогенеза древних Metazoa можно назвать "стомо-фагоцителлой" (подчеркивая эпителизацию лишь ротового отдела фагоцитобласта).
Возможно, на этой стадии филогенеза произошла первая крупная дивергенция филогенетического ствола древних многоклеточных, связанная с тем, что некоторые группы этих животных перешли к освоению морского дна, другие же продолжали совершенствовать приспособления к активной жизни в толще воды.
Современные низшие плоские черви - бескишечные турбеллярии (Acoela) в целом сохранили тот уровень организации, который, вероятно, был характерен для древнейших многоклеточных, впервые перешедших к освоению подвижного образа жизни на дне водоемов. От вендских представителей этих турбеллярий могли возникнуть филогенетические стволы, ведущие к другим группам плоских червей, к нитчатым червям и к предкам кольчатых червей (протоаннелидам). От протоаннелид обособились, с одной стороны, предки моллюсков, с другой - предки членистоногих. У всех этих групп произошла дальнейшая дифференциация фагоцитобласта. У низших червей эпителизировалась лишь его центральная часть, что привело у плоских червей к формированию разветвленного кишечника с единым отверстием - "ртом", ведущим во внешнюю среду, а у нитчатых червей - к образованию сквозного кишечника с ротовым и анальным отверстиями.;У высших групп (кольчатые черви, моллюски и членистоногие) эпителизировался весь фагоцитобласт: не только его центральная часть (энтодермальный кишечник), но и периферическая (мезодерма и ее производные). Последнее привело к развитию вторичной полости тела - целома, стенки которой образованы мезодер-мальным целомическим эпителием. Более примитивные представители кольчатых червей, моллюсков и членистоногих обладают характерной личиночной стадией - трохофорой. В связи с этим указанные группы иногда объединяют под названием Trochozoa.
У тех потомков стомофагоцителлы, которые продолжали совершенствовать адаптации к жизни в толще воды, также произошла эпителизация центрального и отчасти периферического фагоцитобласта: возникла гастральная полость (первичный кишечник) и ее периферические ветви (гастроваскулярные каналы). К этому уровню организации среди современных животных ближе всего стоят гребневики, вероятно сохранившие примитивный образ жизни в толще воды. От их позднепротерозойских предков, которых можно Назвать "проктенофорами", с переходом к прикрепленной жизни На морском дне возникли стрекающие кишечнополостные (тип Coelenterata, или Cnidaria).
Другие филогенетические линии, ответвившиеся от проктенофор, также осваивали морское дно, но с развитием приспособлений к активному передвижению по субстрату, подобно турбелляриям и их потомкам, но на другом исходном уровне организации. У этих форм в результате завершения эпителизации периферического фагоцитобласта также образовалась вторичная полость тела - целом, но возникла она совершенно другим способом, чем у Trochozoa. В онтогенезе животных, происходящих от проктенофор, вторичная полость тела обособляется от первичного кишечника, как его боковые карманообразные выпячивания (первоначально было, вероятно, три пары таких выпячиваний), которые затем отшнуровываются от стенок кишки (рис. 33). Такой способ развития целома получил название энтероцельного - в отличие от схизоиельного способа, характерного для Trochozoa, у которых целом возникает в результате появления полостей внутри скоплений мезодермальных клеток, без всякой связи с первичным кишечником. О. и Р. Гертвиги и И.И.Мечников обосновали гипотезу, согласно которой энтероцельный целом возник в эволюции из гастроваскулярных каналов проктенофорных предков (энтероцельная теория происхождения целома). Энтероцельный целом характерен для типов погонофор (Pogonophora), шетинкочелюстных (Chaeto-gnatha), плеченогих (Brachiopoda), мшанок (Bryozoa) и ряда других, в том числе группы так называемых вторичноротых животных (Deuterostomia), объединяющей типы хордовых (Chordata), иглокожих (Echinodermata) и полухордовых (Hemichordata). У вторичноротых животных имеется много общего, в частности особое положение дефинитивного (присущего взрослым организмам) рта, возникающего на полюсе тела, противоположном первичному эмбриональному рту - бластопору. На месте же последнего развивается анальное отверстие. Вторичноротые, несомненно, имеют общее происхождение; в качестве их предков указывают гипотетическую группу олигомерных червей, тело которых было разделено на три отдела, имелся вторичный рот и энтероцельный целом. Среди современных вторичноротых к уровню организации олигомерных червей ближе всего стоят свободноживущие полухордовые, представителем которых является желудевый червь (Balanoglossus).

Особое положение среди многоклеточных животных занимают губки (тип Porifera, или Spongia). Эта группа характеризуется очень Примитивным общим уровнем организации: губки, по существу, не имеют эпителизованного фагоцитобласта, упорядоченного внутреннего строения, настоящего кишечника, нервной системы, рецепторов и т. д. Губки отличаются от всех остальных Metazoa чрезвычайно своеобразным онтогенезом, в ходе которого происходит инверсия зародышевых листков (экто- и энтодерма, так сказать, меняются местами). Новейшие данные молекулярных исследований показали, что губки имеют общее происхождение со всеми многоклеточными. Вероятно, они представляют собой очень раннюю боковую ветвь, обособившуюся на уровне фагопителлы. Древнейшие ископаемые остатки губок известны из вендских (эдиа-карских) отложений в Австралии.
"Взрывная эволюция" в начале кембрия
Граница между протерозойской и палеозойской эрами отмечается резким изменением в составе и богатстве ископаемой фауны. После в основном "немых" толщ верхнего протерозоя, среди которых рассеяны относительно редкие местонахождения эдиакар-ской фауны, в осадочных породах кембрия (первого периода палеозойской эры), начиная с самых нижних горизонтов, внезапно возникает огромное разнообразие и обилие остатков ископаемых организмов. Среди них помимо низших многоклеточных - губок и кишечнополостных - встречаются также представители высокоразвитых типов животных - плеченогих, моллюсков, членистоногих и др. К концу кембрия появляются почти все известные типы многоклеточных животных. Этот взрыв формообразования на границе протерозоя и палеозоя - одно из самых загадочных событий в истории жизни на Земле. Благодаря этому начало кембрийского периода является столь заметной вехой, что нередко все предшествующее время в геологической истории, т. е. весь криптозой, именуют "докембрием".
Находки эдиакарской фауны и другие данные по докембрийс-ким организмам говорят о том, что эволюция многоклеточных Животных началась задолго до рубежа нижнего кембрия и привела к формированию разнообразных групп. Есть основания считать, что в позднем протерозое уже сложились многие типы Metazoa. Крайняя редкость захоронений ископаемых представителей этих Фупп в отложениях позднего протерозоя, может быть, объясняйся отсутствием у большинства доксмбрийских животных твердого скелета. В раннем кембрии такой скелет появился у самых Разных групп животных. При этом скелетные ткани и анатомия скелета были совершенно различны у разных групп: от гибких членистых хитиновых панцирей членистоногих до монолитных известковых раковин моллюсков и плеченогих. Было высказано предположение, что причиной появления большого количества ископаемых остатков организмов в раннекембрийских отложениях было изменение гидрохимического режима водоемов, благоприятствующее скелетообразованию по чисто химическим причинам, в частности, указывают на усиленное отложение фосфоритов в морских осадках раннего кембрия. Но такое предположение не увязывается с указанным разнообразием форм и химического состава скелета у кембрийских животных. Как могло благоприятствовать повышение содержания фосфатов в воде образованию, например, кремниевой раковины, или хитина, представляющего собой комплекс азотсодержащих полисахаридов? Таким образом, едва ли правильно пытаться свести "нижнекембрийскую революцию" только к скелетизации организмов. Замечательная ископаемая фауна, включающая разнообразных животных, как наделенных скелетом, так и "мягкотелых", была обнаружена в начале 1980-х гг. в средне-кембрийских сланцах Бёрджес в Британской Колумбии в Канаде. В состав этой фауны входит около 120 родов, среди которых представители губок, кишечнополостных, кольчатых червей-полихет, моллюсков, членистоногих, иглокожих, полухордовых, хордовых, плеченогих и других групп, в том числе загадочных организмов неизвестной таксономической принадлежности. Среди них выделяются крупные хищники Anomalocaris, достигавшие 2 м длины и обладавшие странным обликом: удлиненное обтекаемое тело, большие стебельчатые глаза, пара членистых околоротовых придатков, служивших, вероятно, для захвата добычи; челюстной аппарат из множества подвижных пластинок, брюшной плавник, расчлененный на ряд последовательных лопастей. В 1994 г. остатки ископаемой фауны, близкой по составу к фауне Бёрджес, были обнаружены в нижнекембрийских отложениях Южного Китая.
Для объяснения резких изменений состава и общего облика фауны и флоры в некоторые моменты истории Земли, в том числе на рубеже протерозоя и палеозоя, некоторые ученые (в частности, О. Шиндевольф) пытаются привлечь "взрыв мутаций", вызванный какими-то катастрофическими изменениями внешних условий. Подавляющее большинство мутаций снижают жизнеспособность мутантных организмов - этим пытаются объяснить массовое вымирание видов прежней фауны и флоры. В то же время общее усиление мутационного процесса, по мнению Шиндевольфа, приводит к возникновению и быстрому распространению новых форм. При этом сразу возникают новые типы организации - скачкообразно, посредством крупных мутаций. Возможную причину такого "мутационного взрыва" О.Шиндевольф видел в повышении уровня жесткой космической радиации в результате вспышки Сверхновой звезды на достаточно близком расстоянии от Солнца.
К сожалению, эта концепция не может объяснить ни одного конкретного случая изменений фауны и флоры. Почему вымерли одни группы организмов и преуспели другие, прежде сосуществовавшие с первыми? В сущности, гипотеза "мутационного взрыва" просто подменяет анализ конкретных ситуаций постулированием универсальной причины для объяснения любого крупномасштабного эволюционного изменения. Как мы уже неоднократно подчеркивали, основой эволюционного процесса являются малые мутации, и филогенетические преобразования большого масштаба складываются из них под контролем естественного отбора. Крупные же мутации обычно детальны. Наконец, расчеты показали, что вспышки Сверхновых звезд не могут вызвать такого повышения радиации на поверхности Земли, которое имело бы предполагавшийся О.Шиндевольфом мутагенный эффект.
Вообще всегда предпочтительнее искать объяснения событиям, происходящим на Земле, не прибегая к таким гипотезам, которые постулируют универсальное влияние каких-то космических факторов, никак не опосредованное конкретными земными условиями.
В этом отношении от других концепций выгодно отличается гипотеза, выдвинутая в 60-е гг. Л.Беркнером и Л.Маршаллом, объясняющая многие важнейшие события в ходе докембрийской и послекембрийской эволюции, а также на рубеже криптозоя и фанерозоя, опираясь на закономерные изменения условий в среде обитания древних организмов. Эта гипотеза связывает развитие жизни на Земле с изменениями содержания кислорода в земной атмосфере.
В современной атмосфере Земли содержится около 21% кислорода (это соответствует его парциальному давлению 159 мм ртутного столба). Свободный кислород атмосферы необходим подавляющему большинству современных организмов как окислитель в процессах дыхания. Водные организмы обычно дышат кислородом, растворенным в воде, но этот последний образует с атмосферным кислородом единую систему: избыток кислорода, выделяющегося в воде в результате фотосинтеза водных растений, Поступает в атмосферу, а кислород атмосферы растворяется в поверхностном слое воды, по тем или другим причинам обедненной растворенным кислородом.
Однако кислородная атмосфера, столь богатая этим элементом, среди всех планет Солнечной системы присуща только Земле. Это не случайно: высокая химическая активность кислорода приводит к тому, что в условиях планет кислород в свободном состоянии долго существовать не может: участвуя в различных химических реакциях, он оказывается связанным в виде окислов и других соединений. Обилие кислорода в атмосфере современной Земли - результат фотосинтеза, осуществляемого в течение 3 млрд лет зелеными растениями. В процессах фотосинтеза из углекислого газа и воды с использованием энергии солнечного света синтезируются органические вещества (первичная биопродукция) и выделяется свободный кислород.
СО2 + Н2О --> С(Н2О) + О2 + 502 кДж/моль
Первичная атмосфера Земли, существовавшая во времена зарождения жизни, 3,5-4 млрд лет назад, имела восстановительный характер и состояла, вероятно, из водорода, азота, паров воды, углекислого газа, аммиака, метана, аргона и небольших количеств других газов, в том числе и кислорода. Количество свободного кислорода в первичной атмосфере не могло превышать 0,001 от современного его содержания. Это небольшое количество кислорода выделялось в результате фотодиссоциации воды ультрафиолетовыми лучами; кислород быстро входил в различные химические реакции и вновь оказывался в химически связанном состоянии.
Зарождение жизни произошло в бескислородной среде, и кислород из-за своей высокой окислительной способности первоначально был ядовит для протоорганизмов, у которых отсутствовали соответствующие защитные биохимические системы. Вероятно, протоорганизмы по способу питания являлись гетеротрофами, использовавшими в пищу различные органические соединения абиогенного происхождения, которыми, по мнению большинства ученых, были обогащены водоемы раннего археозоя (состояние "первичного бульона", по А.И.Опарину) и на базе которых и возникла сама жизнь. Для освобождения энергии, необходимой в жизненных процессах, первоначально использовалась анаэробная диссимиляция (брожение):
С6Н12О6 --> 2СН3СН2СН + 2СО2 + 210 кДж/моль
С появлением фотосинтеза (первыми фотосинтезируюшими организмами были синезеленые водоросли) в атмосферу стал выделяться кислород.
В условиях бескислородной атмосферы распространение жизни было гораздо более ограниченным, чем ныне. Дело в том. что организмы не имеют эффективных механизмов защиты от гибельной жесткой ультрафиолетовой части излучения Солнца (с длиной волны менее 250 им). В современной атмосфере жесткая ультрафиолетовая радиация поглощается озоновым экраном - слоем озона (О3), образующегося на высоте около 50 км из кислорода (О2) под воздействием солнечного излучения и распределяющегося в основном в 15-60 км от земной поверхности. Озоновый экран надежно защищает живые организмы, которые могут существовать как в водоемах, так и на поверхности суши и в нижних слоях атмосферы. В бескислородной атмосфере раннего протерозоя озоновый экран отсутствовал, и жизнь могла развиваться только под зашитой слоя воды толщиной около 10 м. Получающие наибольшие количества энергии солнечного излучения поверхностные слои воды были недоступны для организмов. Естественно, что совершенно безжизненны были и материки.
Однако фотосинтез, осуществлявшийся в океане планеты синезелеными, а позднее и различными группами эукариотических водорослей, в течение 2 млрд лет медленно, но неуклонно повышал содержание свободного кислорода в атмосфере. Когда содержание кислорода достигло 0,01 от современного (так называемая точка Пастера, соответствующая парциальному давлению кислорода 1,59 мм ртутного столба), у организмов впервые появиласьвозможность использовать для удовлетворения своих энергетических потребностей аэробную диссимиляцию. Другими словами, после достижения точки Пастера стало возможно дыхание, которое почти в 14 раз энергетически эффективнее, чем брожение:
С6Н12О6 + 6О2 --> 6СО2 + 6Н2О + 2,87 мДж/моль - по сравнению с 210 кДж/моль, освобождающимися при брожении. Это был важнейший переломный момент в развитии жизни.
Среди современных организмов так называемые факультативные аэробы, каковыми являются многие бактерии и некоторые дрожжевые грибы, при уменьшении содержания кислорода ниже точки Пастера используют брожение, при повышении его содержания выше этой точки - дыхание (эффект Пастера).
Переход к аэробной диссимиляции в эволюции древних организмов произошел, разумеется, не сразу - для этого необходимо развитие соответствующих ферментативных систем, - но виды, приобретшие способность к дыханию, получили огромный энергетический выигрыш и в результате возможность резко интенсифицировать метаболизм и все жизненные процессы. (как выяснилось, ферментная система клеточного дыхания возникла путем небольшой модификации ферментной системы фотосинтеза - см. обзор по происхождению эукариот; и впрямь, ведь эти два процесса основаны на почти одной и той же последовательности хим. реакций, только идущей в противоположные стороны; при этом надо учитывать, что практически все биохимические реакции обратимы. Это замечательный пример "смены функций" на молекулярном уровне, показывающий к тому же, что даже крупнейшие эволюционные новообразования могут возникать путем небольших модификаций чего-то уже имеющегося в организме - А.М.). Это явилось предпосылкой к дальнейшей прогрессивной эволюции и, вероятно, способствовало ускорению эволюционных преобразований.
Но достижение пастеровской точки в развитии атмосферы Земли Знаменовалось не только появлением возможности аэробной Диссимиляции. При содержании кислорода в атмосфере в количестве 0,01 от современного формируется озоновый экран, который может защитить от жесткой ультрафиолетовой радиации уже и верхние слои воды в водоемах (требуется "помощь" лишь примерно 1 м воды). Это, во-первых, позволяет организмам освоить верхние слои водоемов, наиболее богатые солнечной энергией; в результате резко усиливается эффективность фотосинтеза, увеличивается биопродукция и выделение свободного кислорода. Во-вторых, чрезвычайно расширяется арена жизни: условия в водоемах значительно разнообразнее на малых глубинах, чем на больших. Освоение этого разнообразия условий в богатой энергией среде обитания неминуемо должно было привести к резкому повышению разнообразия форм жизни, к подлинному взрыву формообразования.
По расчетам Беркнера и Маршалла, точка Пастера в эволюции атмосферы Земли была пройдена примерно 620 млн. лет назад; по мнению некоторых других ученых, возможно, значительно раньше - в промежутке 700-1000 млн лет назад. Но, во всяком случае, точка Пастера была пройдена в позднем протерозое, незадолго (в геологическом смысле слова) до рубежа нижнего кембрия. В этом Беркнер и Маршалл видят ключ к решению загадки нижнекембрийского взрыва формообразования в эволюции организмов, который последовал за достижением точки Пастера в атмосфере и логически вытекает из последствий этого события (интенсификация метаболизма, освоение множества новых разнообразных местообитаний, усиление фотосинтеза, возрастание биопродукции, убыстрение эволюции).
После достижения содержания кислорода в атмосфере, равного 0,1 от современного, озоновый экран уже в состоянии полностью защитить организмы от действия жесткой ультрафиолетовой радиации. С этого момента организмы могут начать освоение суши как среды обитания. По расчетам Беркнера и Маршалла, это должно было произойти в конце ордовика (около 420 млн лет назад). Действительно, примерно к этому времени относится появление первых наземных организмов (см. гл. 2). (По мнению ряда других ученых, содержание кислорода в атмосфере соответствующее 10% от современного, было достигнуто уже к началу кембрия, примерно 580 млн лет назад).
Современное содержание кислорода в атмосфере было достигнуто в конце пермского периода.
Гипотеза Беркнера и Маршалла привлекательна не только своей логичностью и последовательностью, но и перспективностью дальнейшего развития этих идей. Как мы уже упоминали, на рубеже нижнего кембрия у самых различных групп организмов развивается твердый скелет, облегчающий их фоссилизацию. Формирование скелета может быть также непосредственным следствием повышения содержания кислорода в атмосфере. Как показали Р. и Е. Раффы, при низком содержании кислорода в окружающей среде размеры тела многоклеточных животных не могли быть большими (вследствие низкого уровня метаболизма и энергетики организмов); газообмен с внешней средой, вероятно, осуществлялся диффузно, через поверхность тела; при этом толщина стенок тела не могла превышать нескольких миллиметров. Для таких организмов не возникало необходимости в опорном внутреннем скелете, а защитные наружные скелетные образования препятствовали бы газообмену. К.Тоув пришел к выводу, что у докембрийских животных в условиях малого содержания кислорода в окружающей среде не могло быть также хорошо развитых соединительнотканных образований, формирующих основу для развития скелета. Прочность соединительнотканных структур основывается на содержании в них белка коллагена, в состав которого входит аминокислота оксипролин. Ее образование возможно только при достаточно высоком содержании кислорода в окружающей среде. Следовательно, при низком содержании кислорода синтез коллагена был биохимически затруднен, и организмы не могли иметь прочных соединительнотканных структур, а поэтому и скелета, и сильно развитой мышечной системы (работа которой эффективна лишь при наличии соответствующих опорных образований).
Д. Роудс и Дж. Морзе исследовали распространение различных животных в современных водоемах с пониженным содержанием кислорода в воде (в Калифорнийском заливе и в Черном море). Была обнаружена отчетливая корреляция между содержанием кислорода в воде и характером донной фауны (бентоса). При содержании кислорода менее 0,1 мл на 1 л воды многоклеточные животные в составе бентоса отсутствуют; при 0,3-1 мл/л встречаются небольшие мягкотелые (бесскелетные) животные, зарывающиеся в ил; наконец, в более поверхностных слоях с содержанием кислорода более 1 мл/л обитают самые разнообразные животные, обладающие известковым скелетом. Эти данные представляют своего рода живую иллюстрацию к концепции Беркнера и Маршалла.
Подведем некоторые итоги. Обособление большинства типов животных произошло, вероятно, в позднем протерозое, 550-800 млн лет назад. Примитивные представители всех групп многоклеточных были небольшими лишенными скелета животными. Продолжавшееся накопление кислорода в атмосфере и увеличение мощности озонового экрана к концу протерозоя позволили животным Увеличить размеры тела. Организмы получили возможность широко расселиться на малых глубинах различных водоемов, что способствовало значительному повышению разнообразия форм животных (вендская фауна).
Однако в позднем венде эта древняя фауна многоклеточных Животных подверглась значительному вымиранию, которое сопровождалось новым существенным уменьшением размеров тела организмов, переживших вымирание. Возможно, этому способствовало чрезвычайно мощное оледенение (по некоторым данным, среднегодовая температура Земли понижалась в это время примерно до 5°С). К концу венда ледники отступили, и условия вновь стали благоприятными для широкого расселения организмов.
(Более подробно о возникновении жизни - см. http://warrax.croco.net/51/eskov/cover_eskov.html).
ГЛАВА 2. ЖИЗНЬ В ПАЛЕОЗОЙСКОЙ ЭРЕ
Итак, в раннем кембрии произошел взрыв формообразовательной эволюции, который в течение примерно 15 млн лет привел к появлению в палеонтологической летописи представителей почти всех известных типов организмов. В течение палеозойской эры, продолжавшейся свыше 300 млн лет, произошли значительные изменения физико-географических условий: рельефа суши и морского дна, общего соотношения площади материков и океанов, положения континентов, климата и многих других факторов. Эти изменения неизбежно должны были сказываться на развитии жизни.
Арена жизни в палеозое
На протяжении палеозоя море неоднократно наступало на континент, заливая опускавшиеся участки континентальных платформ - морские трансгрессии начала кембрия, первой половины ордовика, когда была затоплена наибольшая за весь фанерозой часть континентов, раннего силура, девона и карбона. Морские трансгрессии сменялись поднятием суши, сопровождавшимся горообразовательными процессами и морскими регрессиями (отступление морей) во второй половине кембрия, в конце ордовика, в самом начале и в конце силура, в середине карбона и в пермском периоде. Мощные горные хребты поднялись в конце силура в Скандинавии, Шотландии, Гренландии, на востоке Северной Америки и в ряде других районов земного шара (Каледонская фаза горной складчатости), в каменноугольном и пермском периодах - в Средней Европе (Арденны, Судеты, Гарц, Шварцвальд и др.), на Урале и в Средней Азии (Алай, Тянь-Шань, Ку-энь-Лунь), на севере Сибири, в Австралии и др. (Герцинская фаза складчатости).
Согласно классической теории, которая берет начало в работах Ч.Ляйеля, происходившие в геологической истории изменения границ суши и моря объясняются вертикальными смешениями участков земной коры. Однако ныне все большее признание получает концепция мобилизма -- горизонтальных перемещении континентальных блоков, или "дрейфа континентов", выдвинутая в начале XX в. А. Вегенером. Современный вариант этой концепции - теория тектоники литосферных плит, или "новая глобальная тектоника" - - основывается на глобальном синтезе результатов геолого-геофизических наблюдений и во многом отличается от взглядов Вегенера.
По современным геофизическим данным, астеносфера - верхний слой верхней мантии Земли, располагающийся на глубинах 60-250 км, - имеет пониженную вязкость. Предполагают, что в мантии существуют конвекционные потоки ее вещества, источником энергии для которых являются, вероятно, радиоактивный распад и гравитационная дифференциация вещества мантии. Движение этих потоков со скоростью несколько сантиметров в год происходит в горизонтальной плоскости в верхнем слое верхней мантии от областей подъема к областям опускания ее вещества. В это движение вовлекаются литосферные плиты, как бы плавающие в состоянии изостатического равновесия на поверхности астеносферы и служащие основанием для континентов. При столкновении континентальных плит их края деформируются с возникновением зон складчатости и проявлений магматизма. При столкновении континентальной и океанической плит последняя как бы подминается первой и растекается под ней в астеносфере. Концепция мобилизма в ее современном виде не свободна от противоречий и, вероятно, еще подвергнется существенным перестройкам. Однако уже и теперь она дает удовлетворительное объяснение целому ряду как геологических, так и биологических проблем. К последним относятся, в частности, характерные особенности географического распространения многих видов животных и растений, а также некоторые исторические изменения фауны и флоры.
Согласно представлениям тектоники плит положение и очертания материков и океанов в палеозое были совершенно иными, чем теперь. Палеогеографические реконструкции, выполненные различными учеными с использованием разных методов, довольно существенно различаются в деталях. Однако большинство авторов согласны, что в кембрии южные материки (Южная Америка, Африка, Австралия, Антарктида, а также Индия) были объединены в суперконтинент, называемый Гондваной (рис. 34). Гондвана располагалась главным образом в Южном полушарии, от полюса (находившегося в современной Северо-Западной Аф-Рике) до экватора. В основном в экваториальной зоне находились меньшие по размерам северные материки: Северо-Американский, Восточно-Европейский, Сибирский и микроконтиненты (Средне-Европейский, Казахстанский и др.). Между ними и Гондваной был океан.
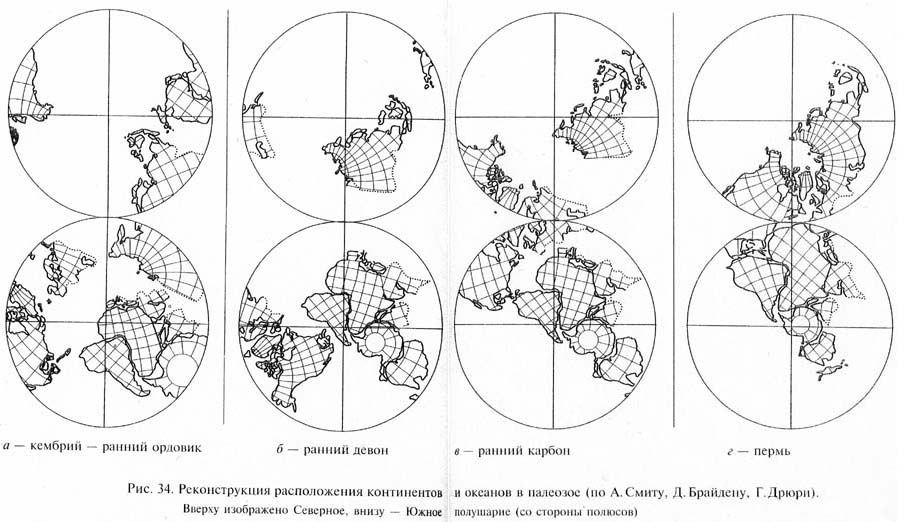
В ордовикском периоде северные континенты начинают постепенно сближаться друг с другом. В силуре и девоне Северо-Амери-канский континент объединился с Восточно-Европейским, с образованием Евроамерики. Пояс Каледонской складчатости в Северо-Западной Европе и Восточной Северной Америке сформировался при столкновении этих континентальных плит. В раннем карбоне сближение континентов продолжалось. Этому соответствовало возникновение поясов Герцинской складчатости. Процесс сближения континентов завершился во второй половине палеозоя (поздний карбон - ранняя пермь) образованием гигантского суперконтинента Пангеи, протянувшегося от Южного полюса, который сместился к этому времени в Антарктиду, далеко в Северное полушарие. Северные материки образовали в составе Пангеи комплекс, получивший название Лавразии.
Перемещения континентальных плит, сопровождавшиеся изменениями рельефа суши и морского дна, обусловливали изменения направлений морских течений и атмосферной циркуляции. Сложные взаимодействия всех этих факторов определяли климатические изменения как местного, так и глобального характера. Согласно Ч.Бруксу, относительное увеличение площади суши в высоких широтах, особенно расположение полюса на суше, вызывает общее понижение температуры поверхности планеты, тогда как увеличение площади суши в низких широтах и расположение полюсов в океане - повышение температуры.
В первой половине палеозойской эры на Земле господствовал в целом теплый климат: средняя температура поверхности Земли была сравнительно высокой, а перепад температур между полюсами и экватором невелик, климатическая зональность была выражена относительно слабо. Теплый период в конце палеозоя, начиная со второй половины карбона, сменился холодным, с более низкой средней температурой, резко выраженной климатической зональностью и значительной разницей температур между полюсами и экватором. Возможно, это связано с расположением в это время центральной части Гондваны в районе Южного полюса. В позднем карбоне и ранней перми мощный ледниковый щит покрывал Антарктиду, Австралию, Индию, южные части Африки и Южной Америки - это было время великого оледенения Гондваны. В верхнепалеозойских отложениях всех этих районов широко распространены тиллиты - ископаемые остатки древних ледниковых морен. В первой половине палеозоя тиллиты имели гораздо более ограниченное распространение; по-видимому, оледенение в конце ордовика и девоне затрагивало лишь приполярные районы (Северо-Западная Африка в ордовике, Аргентина в силуре и девоне). Северный полюс в палеозое находился в океане.
Жизнь в палеозойских морях и пресных водоемах
Вероятно, в кембрийском периоде, как и в докембрии, основные проявления жизни были сосредоточены в морях. Однако организмы уже заселили все разнообразие доступных в море местообитаний, вплоть до прибрежного мелководья, и, возможно, проникли в пресные водоемы.
Водная флора была представлена различными водорослями, основные группы которых возникли еще в протерозое.
Теплые моря кембрийского периода были обильно населены разнообразными животными. Наиболее богата была донная фауна неглубоких морей, прибрежных отмелей, заливов и лагун (во время морских трансгрессий мелководные местообитания получают очень широкое распространение). На морском дне жили разнообразные прикрепленные животные: губки, кишечнополостные (одиночные и колониальные представители различных групп полипов), стебельчатые иглокожие, плеченогие и др. Большинство из них питалось различными микроорганизмами (простейшие, одноклеточные водоросли, бактерии и др.), которых они различными способами отцеживали из воды (фильтрационное питание). Некоторые колониальные организмы, обладавшие известковым
скелетом, возводили на дне моря рифы, подобно современным коралловым полипам. В раннем палеозое это были гидроидные полипы-строматопоры (подкласс Stromatoporoidea), древние коралловые полипы-табуляты (подкласс Tabulata), мшанки (тип Bryozoa) и археоциаты (тип Archaeocyatha). Археоциаты - группа примитивных прикрепленных животных, широко распространенных в раннем кембрии, но полностью вымерших к концу силура. Археоциаты имели известковый скелет в виде своеобразного кубка (рис. 35), стенки которого пронизывали многочисленные поры. Размеры таких кубков варьировали от нескольких миллиметров до 40 сантиметров. Снаружи кубок был одет мягкой оболочкой. Мягкие ткани внутри кубка, видимо, имели губчатое строение, будучи пронизаны системой полостей.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 752; Нарушение авторских прав?; Мы поможем в написании вашей работы!