КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Часть II. Микроэволюция и видообразование 4 страница
|
|
|
|
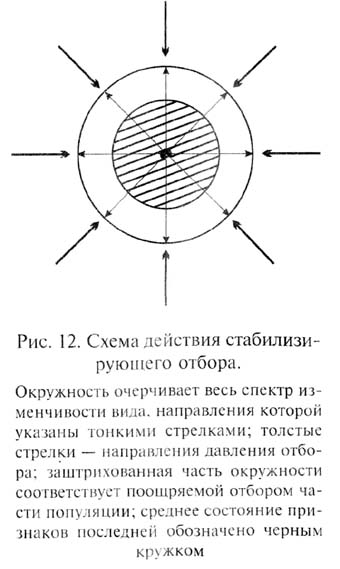
Очевидно, это направление отбора прямо противоположно схеме дивергентной эволюции, разработанной Ч.Дарвином. Под контролем стабилизирующего отбора, действующего длительное время, вид может внешне оставаться неизменным. Однако эта неизменность вида, в сущности, кажущаяся - она касается лини, постоянства оптимальных фенотипов. И.И.Шмальгаузен, разработавший концепцию стабилизирующего отбора, подчеркнул, что в таких условиях генофонд вида продолжает изменяться за счет накопления мутаций, фенотипическое проявление которых взаимно нейтрализуется (важную роль в этом играют гены-модификаторы), а также посредством накопления в гетерозиготном состоянии рецессивных мутаций. Таким образом, генофонд вида обогащается новыми аллелями, которые, оставаясь скрытыми, увеличивают мобилизационный резерв изменчивости, повышающий устойчивость вида к возможным изменениям внешних условий. Эти выводы подкрепляются результатами исследований наследственной изменчивости у разных видов, обитающих в стабильных внешних условиях - например у ряда морских животных (ракообразные, двустворчатые моллюски, иглокожие и другие группы). Оказалось, что генетическая гетерогенность популяций у видов, обитающих в стабильных условиях, не отличается от таковой у видов, обитающих в изменчивой среде.
При действии стабилизирующего отбора может происходить даже дивергенция видов, принимающая, однако, своеобразный характер. Механизм такого видообразования мы рассмотрим в следующей главе.
Направленный отбор наблюдается в том случае, если условия среды благоприятствуют какому-либо одному определенному направлению изменений признака, тогда как все другие варианты изменений подвергаются в той или иной степени негативному давлению отбора (рис. 13). В этом случае в популяции от поколения к поколению происходит сдвиг средней величины признака в определенном направлении. Результатом действия направленного отбора является постепенное изменение популяции в целом, происходящее посредством преобразования популяционного генофонда. В отличие от стабилизирующего отбора изменение частоты встречаемости различных аллелей и закрепление новых мутаций в генофонде при действии направленного отбора быстро отражаются на фенотипическом облике популяции. Это происходит тем быстрее, чем больше интенсивность отбора, если только последняя не превосходит приспособительных возможностей данной популяции - в последнем случае популяция вымирает.
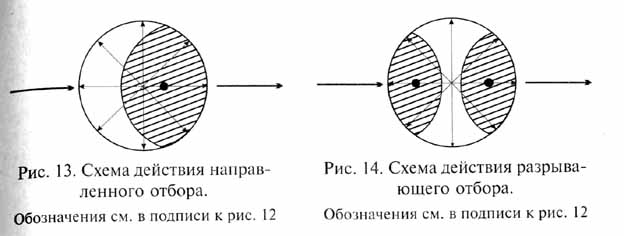
При направленном отборе особенно ярко проявляется воздействие отбора на темпы и преобладающее направление наследственной изменчивости (длящаяся изменчивость, по Ч.Дарвину): получают преимущества такие генотипы, которые обладают соответствующими возможностями дальнейших мутационных изменений, частота определенных мутаций может быть повышена с помощью генов-мутаторов; отбираются гены-модификаторы, способствующие доминантному фенотипическому проявлению благоприятных мутаций.
Осуществляемые под контролем направленного отбора эволюционные преобразования данной популяции затрагивают ее генофонд как целое; обособления каких-либо дочерних популяций (т.е. дивергенции) не происходит. Такую форму эволюционных преобразований вида Д.Симпсон назвал филетической эволюцией.
Разрывающий, или дизруптивный, отбор действует в том случае, когда условия благоприятствуют двум или нескольким крайним вариантам (направлениям) изменчивости, но не благоприятствуют промежуточному, среднему состоянию признака (рис. 14). В результате происходит возрастание изменчивости с последующим сдвигом среднего состояния признака в разных направлениях. Разрывающий отбор ведет либо к возникновению устойчивого адаптивного полиморфизма популяции, либо к ее дроблению, т. е. к разделению на несколько новых популяций такого же ранга. Последний вариант эволюционных изменений всего ближе к дарвиновской концепции дивергентной эволюции.
Тот или другой результат определяется конкретными условиями действия разрывающего отбора: при наличии какой-либо функциональной взаимозависимости между разными благоприятствуемыми генотипами в единой панмиктической популяции возникает полиморфизм, тогда как под влиянием различных вариантов условий в разных частях ареала при снижении скрещивания между населяющими их субпопуляциями происходит дивергенция последних. Д. Симпсон назвал последнюю форму эволюционных изменений видообразованием, поскольку она ведет к возникновению от одного родительского вида двух или нескольких новых - в противоположность филетической эволюции, преобразующей вид как целое, без разделения на дочерние виды.
При действии дизруптивного отбора у молодых дивергирую-щих видов соотношение фенотипической (модификационной) и комбинативной изменчивости становится противоположным тому состоянию, которое возникает под контролем стабилизирующего отбора: системы канализованности индивидуального развития разрушаются, в популяции происходит значительное усиление комбинативной и модификационной изменчивости, что приводит к возрастанию фенотипического разнообразия. Такая картина изменчивости была описана В.В.Кузнецовым в пределах комплекса видов сиговых рыб Coregonus muksun. Быстрое фе-нотипическое выявление разных вариантов комбинативной изменчивости повышает эффективность действия естественного отбора.
И разрывающий, и направленный отбор приводят к векторизованным изменениям генофонда и фенотипического облика популяций; сходно также их воздействие на онтогенез (разрушение систем его канализованности). Отдельные направления разрывающего отбора, действующие на разные аллопатрические популяции одного вида, можно рассматривать применительно к каждой данной популяции как направленный отбор. Поэтому иногда эти две формы отбора объединяют как движущий отбор, противопоставляемый стабилизирующему.
По отношению к разным адаптациям, различным признакам фенотипа у организмов одного вида отбор может иметь разное направление: для одних он оказывается движущим, для других -стабилизирующим. Когда давление движущего отбора по отношению к данной адаптации ослабевает, стабилизирующий отбор способствует закреплению достигнутых движущим отбором результатов, благоприятствуя оптимизации путей морфогенеза у новых адаптивных признаков для достижения максимальной надежности их воспроизведения в фенотипе (с повышением пенетрантно-сти и экспрессивности соответствующих аллелей посредством воздействия генов-модификаторов, а также механизма генокопиро-вания, см. гл. 1).
Одним из ярких примеров действия движущего отбора является развитие так называемого "индустриального меланизма", описанного более чем у 70 видов бабочек, распространенных в Англии и ряде других стран Европы, а также в США. Индустриальный меланизм представляет собой значительное повышение частоты встречаемости меланистических (имеющих темную окраску) особей в тех популяциях бабочек, которые обитают в промышленных районах. Особенно хорошо изучено это явление у березовой пяденицы Biston betularia (рис. 15) Англии.
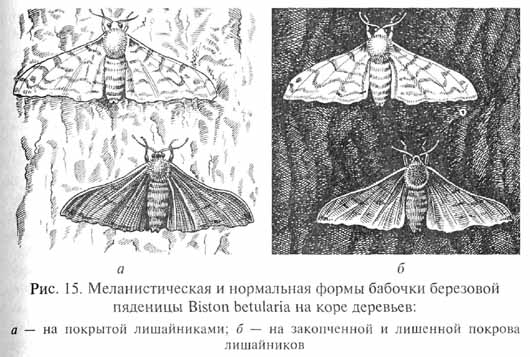
До середины XIX в. в популяциях этого вида преобладали особи, имевшие светлую серовато-белую окраску с темными пятнышками, хорошо маскирующую бабочек на покрытых лишайниками стволах деревьев. В 1848 г. в Манчестере был отловлен первый экземпляр бабочки, имевший меланистическую окраску (морфа carbonaria). С тех пор частота встречаемости меланистических особей в популяциях этой бабочки, обитающих в промышленных районах Англии, постепенно возрастала, пока к началу XX в. количество меланистов не достигло в отдельных популяциях 95% всех особей. Популяции с преобладанием светлоокрашенных бабочек обитают ныне лишь на юго-западе Англии, в Северной Шотландии и в Ирландии. Варианты окраски у березовой пяденицы контролируются аллелями одного гена; аллель, обусловливающий темную окраску, имеет доминантное фенотипичес-кое проявление.
Возможно, меланистические особи в небольшой концентрации встречались в популяциях бабочек и до того момента, когда на них впервые обратили внимание ученые. Однако более широкому распространению аллеля, обусловливающего темную окраску насекомых, в те времена не благоприятствовали условия: ба-бочки-меланисты были гораздо лучше заметны на светлом фоне лишайников, покрывавших кору деревьев, чем "нормальные" светлоокрашенные особи. С развитием промышленности загрязнение воздуха вокруг промышленных центров привело к гибели лишайников на стволах деревьев, которые потемнели от копоти. В этих условиях меланистические формы бабочек оказались в лучших условиях, чем особи со светлой окраской, поскольку первые менее заметны на темных стволах деревьев, чем вторые. Это и определило быстрое увеличение концентрации бабочек-меланистов в популяциях промышленных районов. Благодаря меньшей "задымленности" воздуха в Ирландии, Северной Шотландии и Юго-Западной Англии там сохранилось прежнее преобладание светлой формы В. betularia.
Г. Кеттлуэлл в 50-60-х гг. XX в. экспериментально проверил эти соображения, выпустив определенное количество бабочек темной и светлой окраски в загрязненном лесу вблизи Бирмингема и затем повторив этот опыт в незагрязненном лесу в Дорсете. По данным Кеттлуэлла, птицы, питающиеся бабочками и, так сказать, непосредственно осуществляющие отбор последних, в Бирмингеме уничтожили значительно больше светлых бабочек, чем темных, тогда как в Дорсете наблюдалось обратное соотношение. Явление индустриального меланизма предоставляет ученым замечательные возможности для наблюдений за действием отбора в естественных популяциях. Этот эксперимент, "поставленный" природой при участии человека, уже показал, что изменения генофонда популяций могут происходить под контролем отбора чрезвычайно быстро - в течение нескольких десятилетий. Новые наблюдения показывают, что различия меланистических и светлых особей не сводятся только к разнице в окраске. Кеттлуэлл показал, что и те, и другие бабочки выбирают для отдыха субстраты предпочтительно неконтрастирующей окраски, т.е. ме-ланисты предпочитают темный, а "нормальные" особи - более светлый фон. Имеются также данные о различной жизнеспособности гусениц разных форм бабочек. Возможно, за сто с лишним лет, прошедшие со времени обнаружения первых бабочек-меланистов, не только перестроился генофонд отдельных популяций березовой пяденицы, но и произошли определенные преобразования внутри некоторых генных комплексов с участием генов-модификаторов, которые привели к повышению жизнеспособности и изменениям поведения у темных особей. Эти преобразования могли коснуться и степени доминантности фенотипического проявления того и другого аллелей в пользу повышения доминантности аллеля меланизма. Можно ожидать, что дальнейшее изучение индустриального меланизма и подобных ему явлений позволит проверить и многие другие положения теории отбора.
В обычном понимании отбор представляет собой преимущественное выживание и оставление потомства наиболее приспособленными особями. Можно сказать, что в этом аспекте отбор выступает как своего рода "эгоистический" фактор: видовые адаптации совершенствуются как адаптации отдельных особей. Однако, как мы уже подчеркивали в предшествующей главе, популяции и вид в целом являются целостными системами. Поэтому отбор должен также благоприятствовать закреплению и распространению признаков, которые сами по себе для отдельных особей не имеют приспособительной ценности или даже снижают их приспособленность, но обеспечивают существенные преимущества данной группировке особей как целому. Примеры действия такого "альтруистического" аспекта естественного отбора особенно многочисленны в поведении высших животных. Так возникли "сигналы тревоги", характерные для многих видов птиц: обнаружившая врага птица не затаивается в укрытии (что было бы наиболее благоприятно для ее собственной безопасности), а издает громкий крик, выдающий ее местонахождение хищнику, но предупреждающий других особей своего вида (часто сигнал тревоги воспринимается и другими видами). К этой же форме поведения принадлежит коллективная защита детенышей в стаях и стадах некоторых видов млекопитающих (слоны, копытные), защита и помощь другим особям своего вида (например, поддерживание на плаву раненых животных некоторыми китообразными).
При развитии подобных альтруистических особенностей поведения отбор оперирует не отдельными особями, а целыми их совокупностями (минимальной из них является так называемый дем - группа особей, непосредственно связанных в процессе размножения), благоприятствуя сохранению тех из них, в генофонде которых появляются "альтруистические" аллели. Этот аспект действия естественного отбора получил название "групповой отбор".
Концепция полового отбора была разработана Ч.Дарвином для объяснения развития у самцов некоторых видов животных украшающих признаков, нередко гипертрофированных, причудливых и ярких и поэтому отрицательно сказывающихся на защищенности этих особей и требующих для своего развития значительных энергетических затрат. Очевидно, предпочтение самками самцов с подобными признаками нуждается в объяснении. Первая попытка такого объяснения была сделана Р.Фишером, предположившим, что половой отбор, благоприятствующий развитию украшающих признаков, может действовать на основе предпочтения некоторыми самками таких признаков у самцов. В этом случае самцы, обладающие подобными украшениями, получают лучшие шансы оставить потомство, в котором самцы унаследуют от своих отцов эти признаки, а самки - от своих матерей соответствующие сексуальные предпочтения. Этот механизм (именуемый "фишеровскими процессами") обусловливает возникновение полового отбора, благоприятствующего дальнейшему развитию украшающих признаков у самцов. При этом первоначальное предпочтение самками определенных внешних признаков самцов может быть обусловлено особенностями физиологии их органов чувств (гипотеза первоначального сенсорного предпочтения).
А. Захави предложил интересное объяснение приспособительного значения украшающих признаков: степень их развития в целом должна соответствовать жизнеспособности самцов, поскольку такие "избыточные" структуры без ущерба для организма могут развивать лишь наиболее сильные индивиды с высоким уровнем обмена веществ и энергетических процессов, который контролируется генетически ("принцип гандикапа"). Тем самым степень развития у самцов таких украшений может служить для самок признаком, позволяющим выбрать наиболее жизнеспособных самцов - носителей "оптимальных" генов. Вероятно, развитие украшающих признаков под контролем полового отбора может происходить при совместном действии "фишеровских процессов" и "принципа гандикапа".
Факторы, влияющие на действие отбора
Действие того или иного типа (формы) отбора на каждый конкретный вид организмов определяется особенностями генетической системы данного вида, спецификой его взаимоотношений с внешней средой и характером изменений внешних условий. Очевидно, каждому из трех указанных выше основных направлений отбора благоприятствуют совершенно разные условия: постоянство условий существования - стабилизирующему отбору, изменение условий в одном направлении на всем ареале вида - направленному отбору, разнообразие условий существования (или изменение условий в разных направлениях) в пределах видового ареала - дизруптивному отбору. Вообще в филогенезе всякой группы организмов в течение ее длительной истории направление отбора многократно изменяется, хотя в некоторых случаях долгое время может преобладать какое-либо одно постоянное направление. Длительно действующий направленный отбор приводит к быстрым эволюционным преобразованиям значительного масштаба. Напротив, стабилизирующий отбор в некоторых случаях как бы "консервирует" фенотипический облик отдельных видов организмов в течение длительного времени.
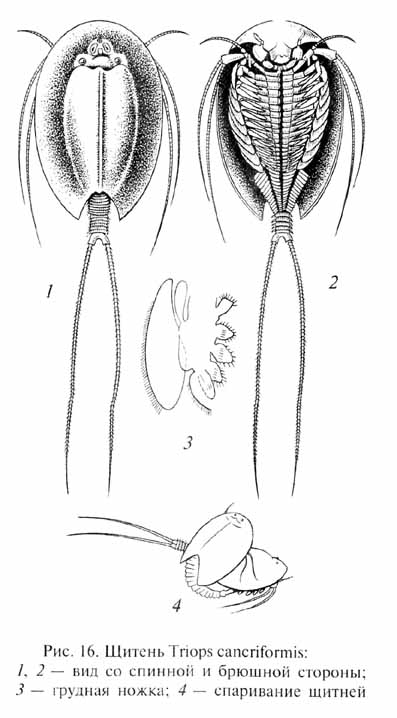
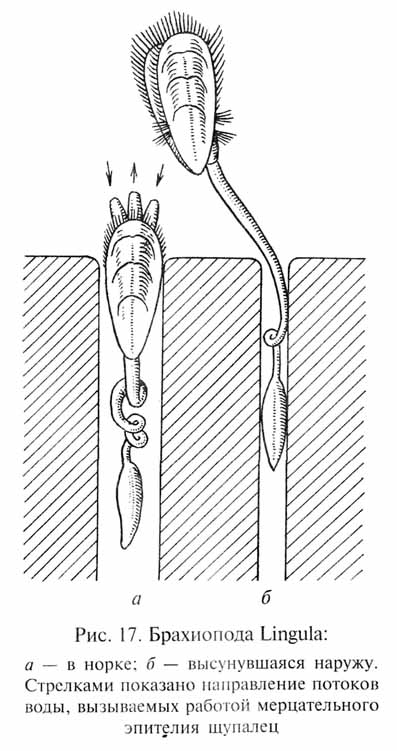
В некоторых местообитаниях абиотические условия относительно мало изменялись или даже оставались в целом стабильными в течение многих миллионов лет (таковы, например, определенные зоны морского дна). В таких условиях некоторые виды организмов могут сохранять свой фенотипический облик практически неизменным в течение значительных периодов времени. Это так называемые персистирующие формы ("живые ископаемые"). Широко известными примерами таких организмов являются равноногие рачки-щитни Triops (рис. 16), организация которых не подвергалась заметным изменениям с триасового периода (более 200 млн лет); двустворчатые моллюски Leda, Nucula, Modiolus, существующие с каменноугольного периода (около 300 млн лет); плеченогие (брахиоподы) Lingula (рис. 17), не претерпевшие изменений с девонского периода(около 380 млн лет). Однако не следует рассматривать персистирующие виды как совершенно те же самые формы организмов,которые жили на Земле сотни миллионов лет назад: как мы уже отмечали, и под контролем стабилизирующего отбора генофонды видов продолжают изменяться, и с течением времени в них накапливается все больше мутаций структурных генов и соответствующи хмутаций генов-модификаторов, препятствующих изменениям фенотипа. Таким образом, в генетическом отношении современные персистирующие виды должны значительно отличаться от своих фЕнотипичЕски сходных предков.
Со времени Ч.Дарвина существенно изменились представления о факторах, благоприятствующих действию естественного отбора. Среди факторов, названных Дарвином (см. гл. 2, ч. I), универсальное значение имеет только длительность времени. Высокая изменчивость благоприятна в наибольшей степени для движущего отбора, стабилизирующий отбор действует против большого размаха изменчивости. Обширность ареала вида благоприятствует дизруптивному отбору.
Географическая изоляция территориальных (аллопатрических) популяций вида ныне рассматривается как один из важнейших факторов, способствующих действию разрывающего отбора. Некоторые ученые считают этот фактор непременным условием процесса видообразования. Под географической изоляцией понимается отделение данной популяции от других популяций родительского вида каким-либо географическим барьером, непроходимым для особей данного вида. Таким барьером может быть морской пролив, горный хребет, русло реки или даже просто район с непригодными для жизни данного вида условиями температуры, влажности, с отсутствием необходимой пищи и т. п.
Численность популяций и дрейф генов
Среди факторов, влияющих на эффективность отбора, особое место занимает численность популяций. С одной стороны, нельзя забывать, что действие отбора вообще имеет вероятностный характер (случайные причины могут привести к гибели или воспрепятствовать участию в размножении отдельных особей, являющихся носителями благоприятствуемых отбором признаков), поэтому для действия отбора необходима достаточно большая численность популяций. Однако, с другой стороны, в очень крупных панмиктических популяциях эффективность отбора может быть в определенной степени снижена благодаря относительной редкости выявления рецессивных мутаций в гомозиготном состоянии; обширный генофонд больших популяций более инертен, чем таковой популяций меньшего масштаба. Поэтому для действия движущего отбора наиболее благоприятны популяции средних размеров, но достаточно большие, чтобы отбор мог оказывать свое влияние вопреки действию случайных факторов.
Численность популяций в природе подвержена значительным колебаниям, зависящим от различных изменений внешней среды (таких, как засуха, морозная или бесснежная зима, урожай или неурожай кормовых растений, численность популяций других видов животных и т. п.). В урожайные годы численность популяций данного вида может существенно повыситься, вслед за чем при ухудшении условий наступает ее спад, тем более резкий, чем больше контраст условий в последовательные годы. Особенно ярко эти "популяционные волны" проявляются у видов с коротким жизненным циклом и быстрой сменой поколений (мелкие грызуны, ящерицы, лягушки, насекомые, однолетние растения, микроорганизмы и т.п.), но колебания численности популяций присущи всем без исключения видам организмов.
С.С.Четвериков подчеркнул большое эволюционное значение колебаний численности природных популяций, которые он назвал "волнами жизни". Поскольку для действия отбора важна численность популяций, очевидно, что ее изменения должны сопровождаться изменениями интенсивности, а в некоторых случаях - и направления отбора. Кроме того, колебания численности популяций неизбежно приводят к изменениям концентраций различных аллелей в популяционном генофонде, которые могут иметь случайный характер (исчезновение редких аллелей при снижении численности популяций или, наоборот, их значительное распространение в генофонде и т. п.). Такие изменения генофонда сами по себе представляют материал для действия отбора (отбор оптимального генофонда при последующей стабилизации условий).
В небольших популяциях, подвергшихся изоляции от других Популяций своего вида, влияние случайных факторов может выйти на первый план по отношению к действию отбора. Эта интересная ситуация в 1931 г. была описана Н.П.Дубининым и Д.Д.Ромашовым под названием "генетико-автоматические процессы" и, независимо от них, в том же году С. Райтом, который обозначил ее термином "дрейф генов". Если небольшая популяция (менее 50 особей) оказывается изолированной, прежде всего проявляется эффект, названный "принципом основателя". Генофонд малочисленной популяции с самого начала беднее, чем генофонд родительского вида, и подбор аллелей в генофонде "популяции основателей" случаен из-за случайного подбора особей. Такой генофонд не сбалансирован, поскольку он возник на основе случайности, а не под контролем естественного отбора. Так как отбор действует на вероятностном уровне и требует для своей эффективности достаточной численности особей, дальнейшая судьба генофонда маленькой популяции определяется в первую очередь действием различного рода случайных факторов.
Общей тенденцией будет выпадение из генофонда малой популяции наиболее редких аллелей, сопровождающееся их замещением аллелями, которые с самого начала были более многочисленными. Это ведет к дальнейшему обеднению генофонда малой популяции и в результате к его гомозиготизации. Важно подчеркнуть, что указанные изменения генофонда не имеют приспособительного характера: утраченные аллели могут обладать равной и даже большей приспособительной ценностью, чем аллели, ставшие в популяции преобладающими: ведь потеря первых определяется не действием отбора, а случайной гибелью особей. Такая случайная гибель особей от самых различных причин (хищники, заболевания, факторы абиотической среды) происходит всегда и в любой популяции, но при большом количестве особей она отступает на задний план - в среднем успешнее выживают и оставляют потомство лучше приспособленные организмы. В малой же популяции фактор случайности становится основным. Естественно, что в этом случае лучшие шансы для сохранения имеют первоначально более многочисленные аллели вне зависимости от их приспособительной ценности.
Таким образом, действие случайных факторов обедняет и значительно изменяет генофонд малой изолированной популяции по сравнению с его исходным состоянием. Это явление и называется собственно генетико-автоматическими процессами или дрейфом генов. В результате подобных процессов может сложиться жизнеспособная популяция со своеобразным генофондом (случайно, поскольку отбор в данном случае не играл ведущей роли!). Спектр изменчивости этой популяции существенно отличается от такового родительского вида. С постепенным увеличением численности особей вновь восстановится эффективность естественного отбора, который будет действовать уже на новый генофонд, обладающий специфическим спектром изменчивости. Совокупность всех этих процессов может привести к обособлению нового вида.
Соотношения роли отбора и дрейфа генов при разной численности популяций можно выразить по С.Райту следующим образом:
если численность популяции N <1/(2 S) (где S - коэффициент интенсивности отбора, см. выше), преобладает воздействие дрейфа генов; если N >1/(4 S) преобладает отбор; в диапазоне N от 1/(4 S) до 1/(2 S) совместно действуют оба фактора. Так, при умеренной интенсивности отбора (S =0,01) изменения соотношений любой пары аллелей регулируются дрейфом генов при /V<50; при слабой интенсивности отбора (S = 0,001) дрейф генов оказывается определяющим фактором при N < 500 и т.д.
В соответствии с этим генетико-автоматические процессы могут существенно влиять на генофонд популяции основателей в первое время после ее закрепления в новом районе, пока она еще достаточно малочисленна; затем на первый план неизбежно выходит действие естественного отбора. Если условия в новом районе останутся благоприятными, численность популяции быстро увеличится и эффект случайности - дрейф генов - будет все более затушевываться влиянием отбора, как бы ни было слабо его давление. Если же условия среды станут неблагоприятными и численность популяции вследствие этого останется небольшой - это явится непосредственным результатом жесткого давления отбора и дрейф опять-таки не сможет быть определяющим фактором эволюционных перестроек генофонда в течение длительного времени.
Сравнивая эволюционное значение рассмотренных нами факторов, мы можем заключить, что наличие мутаций и естественного отбора необходимо и достаточно, чтобы обеспечить приспособительную и дивергентную эволюцию организмов. Поэтому мутационный процесс и естественный отбор можно обозначить как необходимые факторы эволюционного процесса. При этом отбор является единственным известным ныне фактором эволюции, который объединяет движущее, направляющее и интегрирующее воздействия на организмы, формируя их приспособления и влияя на саму мутационную изменчивость. Прочие рассмотренные нами элементарные эволюционные факторы (колебания численности, обмен генетической информацией между разными популяциями, географическая изоляция, дрейф генов) являются дополнительными по отношению к мутационному процессу и естественному отбору. Разумеется, для полного понимания эволюционного процесса необходимо учитывать сложные взаимодействия всех указанных элементарных эволюционных факторов.
ГЛАВА 4. ВИДООБРАЗОВАНИЕ И МЕЖВИДОВЫЕ ОТНОШЕНИЯ
Среди различных результатов эволюционного процесса видообразование в наибольшей мере привлекало внимание исследователей на всех этапах развития эволюционистики (не случайно Ч.Дарвин назвал свой основной труд "Происхождение видов"). Именно этот аспект эволюционного процесса стал основным содержанием синтетической теории эволюции, которая, в сущности, представляет собой главным образом теорию видообразования. Проблема видообразования находится в центре внимания и некоторых альтернативных эволюционных концепций (сальтационизма, пунктуализма).
Аллопатрическое видообразование
Как мы уже упоминали, всякая территориальная популяция характеризуется собственным генофондом, частота встречаемости разных аллелей в котором оптимизирована отбором применительно к условиям обитания данной популяции. При обособлении популяции обмен наследственной информацией ("поток генов") между нею и остальными популяциями прекращается. Генофонд изолированной популяции становится самостоятельным. Особенности местных условий определяют специфическое направление отбора, действующего на такую популяцию (это направление является одним из направлений дизруптивного отбора, действующего на вид в целом). Постепенно в генофонде изолированной популяции происходит накопление новых мутаций. Поскольку генофонд данной популяции специфичен, спектр его изменчивости должен несколько отличаться от такового других популяций вида. Специфика местных условий и соответствующее направление отбора усугубляют особенности генофонда изолированной популяции. Поэтому мутации, накапливающиеся в ее генофонде, будут в той или иной степени отличаться от мутаций, которые за это же время закрепятся в других популяциях родительского вида.
Если географическая изоляция достаточно продолжительна, а направления отбора достаточно различны, то изолированная популяция приобретает отчетливые различия с родительским видом, снижающие возможность скрещивания, вплоть до возникновения репродуктивной изоляции (т.е. нескрещиваемости в природных условиях) между ними. Репродуктивная изоляция может основываться на самых разных биологических особенностях близких форм организмов. Выделяют следующие основные формы репродуктивной изоляции: 1) этологическая (различия поведения, особенно брачного); 2) экологическая (различия предпочитаемых местообитаний); 3) временная, или сезонная (различия сроков размножения); 4) морфологическая (различия размеров тела и особенностей строения, важных для опознания половых партнеров, их взаимной стимуляции и спаривания); 5) генетическая (различия генома, приводящие к генетической несовместимости половых клеток). Все эти и другие неупомянутые здесь формы изоляции возникают независимо друг от друга и могут сочетаться в любых комбинациях или действовать поодиночке. Наиболее совершенна генетическая изоляция, исключающая всякую возможность обмена генетической информацией между двумя популяциями.
После возникновения той или иной формы репродуктивной изоляции обособленная популяция становится самостоятельным видом. Если теперь географический барьер, разделявший популяции, нарушится и вновь восстановится контакт между ними -эти два вида либо смогут сосуществовать в одном ареале, либо между ними возникнут конкурентные отношения и один из них будет вытеснять другой, если их биология осталась близкой. Но, так или иначе, это будут уже две обособленные филетические линии, каждая из которых имеет свою эволюционную судьбу.
Такая форма видообразования, основанная на территориальной изоляции разных популяций, получила название аллопатрической. Ч.Дарвин также указывал, что географическая изоляция способствует видообразованию, но придавал этому фактору меньшее значение, чем современная теория эволюции.
Если две изолированные друг от друга популяции одного вида находятся в сходных условиях, длительное время остающихся постоянными, на них будет действовать одинаково направленный стабилизирующий отбор. В таких популяциях формируются практически одинаковые оптимальные фенотипы. Однако под прикрытием этих оптимальных фенотипов в генофонде популяций, как мы видели выше (гл. 3), происходит постепенное накопление новых мутаций. Если обмен генетической информацией между двумя такими популяциями затруднен в течение достаточно длительного времени, в их генофондах могут накапливаться разные мутации. В результате популяции, оставаясь внешне сходными, будут все более различаться в генетическом отношении. С накоплением этих различий возможности их гибридизации постепенно снижаются, вплоть до полной невозможности скрещивания (возникновение генетической несовместимости) особей из разных популяций. На этой стадии две обособленные популяции превращаются в два самостоятельных вида, которые, однако, сохраняют фенотипическое сходство. Такие виды носят название видов-двойников. Сходство видов-двойников может быть столь полным, что даже опытные зоологи-систематики нередко с большим трудом различают подобные формы, единственным надежным критерием для разделения которых является их нескрещиваемость друг с другом.
Примером видов-двойников могут служить хорошо изученные формы плодовых мушек Drosophila pseudoobscura и D. persimilis. За 35 лет исследований в природных популяциях было обнаружено лишь 4 достоверно установленных случая гибридизации между этими фенотипически чрезвычайно сходными видами. Обмену генетической информацией между D. pseudoobscura и D. persimilis препятствуют различия поведения (этологическая изоляция), стерильность гибридных самцов, пониженная жизнеспособность и стерильность потомства от возвратных скрещиваний гибридных самок. Виды-двойники обнаружены в большинстве групп животных и растений.
Изолированные популяции могут возникать двумя путями: либо при появлении каких-либо географических барьеров, разделяющих прежний ареал вида, либо при активном расселении вида на новые территории за пределы занимаемого им ареала. В этом случае относительно небольшие популяции, укоренившиеся в новых районах, могут с самого момента своего возникновения стать территориально обособленными (периферические изоляты). Этот последний случай особенно интересен тем, что периферические изоляты могут сразу подвергнуться действию условий, существенно отличающихся от таковых в пределах ареала вида. Э.Майр высказал предположение, что в периферических изолятах могут в некоторых случаях происходить относительно очень быстрые и резкие изменения генофонда, приводящие к эволюционным изменениям значительного масштаба. Этот процесс получил название " генетическая революция ". Основную причину генетической революции Э.Майр видит в том, что генофонд периферического изолята (или популяции основателей) с самого начала не сбалансирован и не оптимизирован отбором. Поэтому при закреплении популяции основателей в новом для вида районе ее генофонд должен подвергнуться существенной перестройке под давлением отбора, до установления новой формы генетического равновесия. При малочисленности популяций основателей преобразования их генофондов ускоряются явлениями дрейфа генов.
Г.Льюис подкрепил гипотезу Э.Майра о возможности генетических революций анализом последствий действия на такие популяции особенно интенсивного отбора, названного им " катастрофическим ". Популяции, обитающие в краевой зоне ареала вида, часто подвергаются действию необычных для данного вида экстремальных условий. При крайне неблагоприятных условиях на популяцию действует катастрофический отбор, элиминирующий большинство составляющих ее особей. При этом выживают немногие индивиды, обладающие наиболее высокой фенотипической устойчивостью к неблагоприятным факторам (на основе соответствующих генетических уклонений). После возвращения условий к норме численность популяции восстанавливается, но ее генофонд после действия катастрофического отбора должен существенно отличаться от исходного.
Проблема симпатрического видообразования
Некоторые ученые допускают возможность дивергенции видов из одной и той же территориальной популяции, без ее разделения на аллопатрические группировки (т.е. без географической изоляции). Симпатрическое видообразование должно основываться на высокой степени популяционного полиморфизма, причем устойчивые морфы, имеющиеся в одной популяции, не должны быть взаимозависимы. При наличии такого полиморфизма в исходной популяции действие достаточно интенсивного разрывающего отбора, благоприятствующего наиболее уклонившимся морфам и элиминирующего все промежуточные состояния, может привести к снижению панмиксии и затем к обособлению устойчивых биологических рас - симпатрических внутривидовых группировок, различающихся определенными экологическими особенностями. Дальнейшая экологическая специализация биологических рас под давлением интенсивного разрывающего отбора может привести к их репродуктивной изоляции друг от друга. Д.Смит (1966) сформулировал ряд условий, необходимых для успеха симпатрического видообразования под действием дизруптив-ного отбора: 1) обитание вида более чем в одной экологической нише (т.е. использование различными симпатрическими группировками разных местообитаний, разных кормов и способов их добывания и т. п.); 2) отдельное регулирование численности популяций, занимающих каждую нишу; 3) наличие отмечаемых отбором преимуществ каждой популяции в своей нише; 4) возникновение репродуктивной изоляции между указанными симпатрическими популяциями.
Основной трудностью, стоящей перед концепцией симпатрического видообразования, является неизбежность скрещивания в Исходной полиморфной популяции на первых этапах дивергенции биологических рас, которое должно препятствовать обособлению подобных группировок. Крайний скепсис в отношении симпатрического видообразования проявляет Э. Майр, по мнению Которого интенсивность разрывающего отбора, которая необходима для симпатрического видообразования, должна значительно превышать реально наблюдаемую в природе.
Однако в пользу его реальности говорит формирование у некоторых видов устойчивых симпатрических биологических рас. Примером таких группировок могут служить питающиеся разными кормовыми растениями ивовая и березовая расы жуков-листоедов Lochmaea capreae, между которыми, по данным С.Г.Креславского, существует высокий уровень репродуктивной изоляции (97 %).
Существует еще несколько особых форм симпатрического видообразования, обеспечивающих чрезвычайно быстрое обособление дочерней формы от родительского вида: полиплоидизация, гибридогенез (возникновение в результате гибридизации аллополиплоидных форм, см. гл. 1) и так называемый стасигенез (или стасипатрическое видообразование). Последним термином М.Уайт обозначил обособление новых видов в результате крупномасштабных хромосомных мутаций (инверсий, транслокаций). Во всех этих случаях новый вид сразу оказывается генетически изолированным от родительского, обитая на той же самой территории, и сохраняет свою обособленность, если является жизнеспособным в данных условиях и способен к бесполому или партеногенетиче-скому размножению (в связи с этим такие формы видообразования более обычны у растений, чем у животных). Позднее, после накопления достаточного количества особей с однотипным геномом, может восстановиться половой процесс.
Таким образом, формы видообразования достаточно разнообразны, поскольку в рамки двух его основных категорий - аллопатрического и симпатрического - включен, с некоторой долей условности, ряд особых моделей видообразования, существенно отличающихся своими механизмами (так, симпатрическое видообразование включает гибридогенез, стасигенез, полиплоидизацию и дивергенцию биологических рас).
Темпы видообразования
Подобно формам видообразования столь же разнообразны и его темпы, варьирующие от относительно медленного и постепенного преобразования генофондов географически обособленных популяций при "классической" аллопатрической модели до чрезвычайно быстрого (в процессе смены всего нескольких поколений) обособления генофонда популяции основателей при "генетической революции" и действии катастрофического отбора, или столь же быстрого формирования нового вида при полиплои-дизации и стасигенезе.
В 1972 г. Н.Элдридж и С.Гулд выдвинули получившую широкую известность гипотезу так называемого "прерывистого равновесия", или "пунктуализма", которую они считают альтернативной классическим представлениям дарвинизма о постепенном ("градуалистском") видообразовании. Согласно этой гипотезе история любой филетической линии состоит из чередования длительных периодов эволюционной стабильности - до 99% времени существования вида - - и относительно кратких периодов видообразования: каждый акт видообразования охватывает около 10000-100000 лет. В качестве основного аргумента в пользу своей гипотезы Гулд и Элдридж использовали широко распространенное явление прерывистости филетических линий в палеонтологической летописи, обычно объясняемое фрагментарностью палеонтологических данных и приводящее к описанию дискретных во времени палеонтологических видов.
Сторонники пунктуализма видят основные механизмы ускоренного видообразования в процессах генетических революций, происходящих в периферических изолятах; в сериях макромутаций (особенно в мутациях генов-регуляторов и хромосомных перестройках) и в так называемом "видовом отборе". Этот термин используется для обозначения отбора по признакам, инвариантным (единообразным) для вида в целом. В этом случае виды рассматривают как аналоги особей, причем сторонники пунктуализма полагают, что "видовой отбор" должен приводить к быстрому замещению хуже приспособленных по данному признаку видов лучше приспособленными. Однако необходимо помнить, что отбор - это вероятностный процесс, эффективный лишь при достаточно больших выборках. Число конкурирующих видов обычно значительно меньше минимального числа особей в популяции, начиная с которого отбор становится более эффективным, чем дрейф генов. Отбор по инвариантным видовым признакам, конечно, может происходить, но он базируется не на видах как целостных единицах, а на большом числе составляющих их особей. Поэтому такой отбор ничем принципиально не отличается от обычного естественного отбора и не требует выделения в особую категорию.
Проверка многочисленных палеонтологических данных о характере видообразования в различных филетических линиях, проделанная целым рядом ученых, показала, что в палеонтологических рядах форм можно обнаружить и "градуалистское", и "пунктуалистское" видообразование. Вообще же, "разрешающая способность" палеонтологической летописи, как правило, совершенно недостаточна для того, чтобы по палеонтологическим данным можно было с достаточной достоверностью судить о характере процессов видообразования, поскольку "разрывы" между предками и потомками могут быть связаны как с высокими темпами видообразования, так и с неполнотой палеонтологической летописи.
Ускоренное видообразование, утверждаемое сторонниками пунктуализма, вообще говоря, вполне возможно, и механизмы его известны (это "генетические революции", стасигенез, поли-плоидизация, гибридогенез). Однако неверно главное положение гипотезы прерывистого равновесия, что это - единственная (или резко преобладающая) форма эволюции, чему противоречат многочисленные данные по современным и ископаемым формам.
Хотя сама концепция пунктуализма в целом не выдержала критики, дискуссии с ее сторонниками в 70-80-е гг. способствовали развитию представлений о многообразии форм и темпов видообразования.
Взаимоотношения близких видов
После того как между родственными популяциями сформировались какие-либо механизмы репродуктивной изоляции, эти популяции можно рассматривать как самостоятельные виды. При возникновении вторичного контакта и перекрывания ареалов таких форм (неосимпатрия), между ними могут сложиться различные взаимоотношения в зависимости от степени их экологической дивергенции и генетической изоляции.
Гибридизация. Если экологическая дивергенция уже столь велика, что близкие виды не вступают в остроконкурентные отношения, то степень генетической изоляции не является для них определяющим фактором: даже если виды могут скрещиваться, давая плодовитое потомство, полного воссоединения их генофондов обычно не происходит. Пример подобной ситуации был описан у ящериц-игуанид Sceloporus woodi и S. undulatus, ареалы которых перекрываются во Флориде. Хотя этологические механизмы, препятствующие спариванию представителей этих двух видов, отсутствуют и их гибриды плодовиты, зона гибридизации остается очень узкой, не превышая нескольких миль в ширину. Это связано с предпочтениями разных биотопов, использованием разных способов охоты и разных наборов кормов у двух указанных видов ящериц.
Подобная ситуация может, вероятно, сохраняться довольно долго, но при изменениях условий внешней среды в зоне контакта таких видов экологическая изоляция может оказаться малоэффективной, что приведет к усилению гибридизации родственных форм. Подобные ситуации уже многократно описаны в самых различных группах животных. Например, по данным Д.Джонса, в районе Блумингтона (Индиана, США) происходит регулярная гибридизация двух видов жаб Bufo americana и В. woodhousei fowleri.
К настоящему времени в результате гибридизации произошло морфологическое сближение обитающих здесь популяций обоих видов. Предполагают, что причиной гибридизации, до 1941 г. не отмечавшейся и, очевидно, усилившейся вторично, являются изменения внешней среды деятельностью человека (расчистка лесов, загрязнение водоемов).
Интересный пример усиления гибридизации между родственными видами описан в 1973 г. Л.С.Рябовым: в Воронежской области за последние 13 лет отмечено свыше 100 особей-гибридов волка и домашней собаки, причем наблюдается тенденция к увеличению числа этих гибридов и расширению зоны гибридизации. Известно, что обычные отношения волка и собаки в целом антагонистичны - волки нередко нападают на собак, используя их как добычу. Изменения взаимоотношений этих двух видов, по мнению Л. С. Рябова, связаны с сокращением численности волков и нарушением нормального соотношения числа самцов и самок в их популяциях.
В любом случае, если гибридизация двух видов происходит регулярно хотя бы в небольшой зоне и гибриды жизнеспособны и плодовиты, это должно приводить благодаря возвратному скрещиванию к проникновению некоторого количества генов одного вида в генофонд другого. Это явление получило название интрогрессивной гибридизации (или интрогрессии). Интрогрессивная гибридизация может способствовать усилению наследственной изменчивости родственных видов, обогащая их генофонды новыми аллелями. Э. Майр полагает, что при этом возможен и другой эффект -- проникновение путем интрогрессии новых аллелей в уже сложившиеся, сбалансированные и оптимизированные отбором генофонды популяций родственных видов может нарушить сбалансированность этих генофондов, способствуя снижению приспособленности отдельных особей и популяции в целом. В этом случае отбор должен действовать в пользу усиления механизмов репродуктивной изоляции, препятствующих гибридизации. Происходит завершение процесса дивергенции видов, начавшейся в условиях географической изоляции. Такой вариант видообразования, начавшегося как аллопатрическое и завершающегося в условиях неосимпатрии, иногда называют "парапатрическим".
При более отдаленном родстве скрещивающихся видов их генотипы уже настолько различны, что гомология хромосом во многом нарушена. У таких гибридов мейоз затруднен из-за невозможности нормальной конъюгации хромосом в профазе первого Деления, и гаметогенез протекает с существенными нарушения-Ми, так что жизнеспособными оказываются относительно немногие гаметы, в частности получившие полные (двойные) наборы Хромосом без какой-либо редукции их числа. При копуляции таких гамет во втором гибридном поколении возникают так называемые амфидиплоыды - организмы, имеющие диплоидные наборы хромосом от каждого из двух родительских видов. Цитологически амфидиплоид ведет себя как нормальная диплоидная особь, поскольку каждая хромосома приобрела себе гомолога. Поэтому ам-фидиплоиды второго гибридного поколения могут дать начало новому виду, отличающемуся от обоих родительских - если, конечно, эти амфидишюидные организмы окажутся жизнеспособными и смогут выдержать конкуренцию с обоими родительскими видами. Роль такого способа видообразования, по-видимому, сравнительно невелика, хотя в селекции этот способ используется для выведения новых форм сельскохозяйственных растений (см. с. 57).
Если же происходит обратное скрещивание гибридов первого поколения, полученных от подобной отдаленной гибридизации, при копуляции их диплоидных гамет с гаплоидными гаметами родительских видов получаются триплоидные особи, которые также могут дать начало новым формам организмов при условии их способности к вегетативному размножению (у растений), партеногенезу или гиногенезу (у животных). Таков был путь возникновения партеновидов у некоторых групп позвоночных. В частности, подобные партеновиды описаны в двух семействах ящериц - Lacertidae и Teiidae.
Иногда при отдаленной гибридизации возникают своеобразные комплексы взаимосвязанных форм, подобные комплексу бисексуальных диплоидных видов саламандр Ambystoma laterale и A. jeffersonianum с триплоидными самками, возникшими в результате их гибридизации ("A. platyneum") и размножающимися путем гиногенеза (с активацией триплоидных яиц к дроблению гаплоидными спермиями родительских видов). Еще один характерный пример подобных взаимоотношений представляет комплекс видов европейских зеленых лягушек: озерной (Rana ridibunda), прудовой (R. lessonae) и съедобной (R. esculenta). Как показал Л. Бергер, последняя форма возникла в результате гибридизации двух первых видов. Геном R. esculenta включает гаплоидные геномы R. ridibunda и R. lessonae; гаметы съедобной лягушки получают либо оба этих генома, либо лишь гаплоидный геном R. ridibunda. Популяции съедобной лягушки поддерживаются за счет скрещивания этой формы с прудовой лягушкой, а также за счет новых скрещиваний прудовых и озерных лягушек.
Очевидно, такие формы организмов, которые воспроизводятся только путем гиногенеза (Ambystoma platyneum) или гибридо-генеза (Rana esculenta), не являются настоящими видами; для их обозначения введен особый термин - "клептоны".
Конкуренция. При возникновении неосимпатрии близкородственных видов столь же часто возникает и ситуация, противоположная рассмотренной нами выше, - а именно, когда генетическая изоляция двух видов достаточно совершенна и гибридизация -не играет заметной роли, но экологическая дивергенция незначительна. В этом случае два вида имеют весьма сходные экологические потребности (местообитания, корма, убежища, места размножения и т. п.). В результате между близкородственными видами возникают остроконкурентные отношения, причем один вид нередко вытесняет другой. Этот аспект взаимоотношений между близкими видами был рассмотрен Ч.Дарвином в схеме дивергентной эволюции. В соответствии с выводами Дарвина отбор в данном случае направлен на увеличение экологических различий между родственными видами.
Собрано много примеров остроконкурентных, антагонистических взаимоотношений между близкими видами. Так, лягушка-бык (Rana catesbeiana), обитающая в восточной части Северной Америки и завезенная в Калифорнию в 1914-1920 гг., полностью вытеснила из большинства районов Калифорнии два туземных вида лягушек - R. aurora и R. boylii, ныне сохранившихся лишь в таких местообитаниях, которые не заняты лягушкой-быком. На мелких островках в Адриатическом море у побережья Далмации встречаются два вида ящериц - Lacerta melisellensis и L. sicula, причем распространение этих форм, как правило, имеет аллопатричес-кий характер. По-видимому, L. sicula начала колонизовать острова позднее, чем L. melisellensis, и, по данным М.Радовановича, постепенно вытесняет последний вид. Остроконкурентные отношения были описаны у двух видов саламандр-плетодонтид Plethodon cinereus и P. nettingi в восточной части Северной Америки: первый вид вытесняет второй из всех благоприятных местообитаний на более сухие скальные осыпи, будучи более крупным и выигрывая пищевую конкуренцию.
Однако в некоторых случаях экологическая конкуренция близких видов может и не приводить к обязательному вытеснению (конкурентному исключению) одного вида другим. Если лимитирующие ресурсы (т. е. те факторы внешней среды, которые служат основным объектом конкуренции, - пища, оптимальные местообитания, места размножения и т.п.) имеются в районе симпат-рии конкурирующих видов в достаточном количестве, то популяции этих видов могут сосуществовать в одном местообитании в определенном равновесии без особого обострения конкуренции между ними. Это, в частности, может быть обусловлено действием внутрипопуляционных механизмов регуляции численности вида-доминанта, которые ограничивают последнюю более низким уровнем, чем это допускается наличными ресурсами среды. В этом случае неиспользованные ресурсы позволяют обитать на этой же территории также популяциям более слабого в конкурентном отношении вида. Такая ситуация была описана на примере совместного обитания экологически близких видов лягушек Rana blythi и R. ibanorum в одних и тех же местообитаниях в Сараваке (на острове Борнео). Конкуренция между симпатрическими популяциями этих видов лягушек ограничивает их максимальную численность. Однако рост плотности популяций каждого вида не идет дальше определенного предела даже при низкой численности популяции вида-конкурента. Это связано с тем, что возрастание плотности популяции ограничивает собственный рост в большей степени, чем рост популяции вида-конкурента.
Конкурирующие виды могут сосуществовать в одном местообитании и в том случае, когда каждый из них имеет определенные преимущества в разные сезоны года (подобно разным морфам при видовом полиморфизме, см. гл. 2). Так, по данным А.Дондта, большая синица (Parus major) и синица-лазоревка (P. caeruleus) конкурируют за пищу и ночные убежища. В сезон размножения лазоревка выигрывает пищевую конкуренцию, потребляя более мелких гусениц и личинок и лишая большую синицу значительной части пищевых ресурсов. С другой стороны, в остальное время года большая синица имеет преимущество в конкуренции за убежища для ночевки. В результате между симпатрическими популяциями обоих видов устанавливается определенное равновесие.
Межвидовые отношения в биоценозах и коэволюция
Для понимания процессов видообразования и дальнейшей эволюционной судьбы видов организмов необходимо принимать во внимание не только взаимоотношения близких недавно дивергировавших форм, но и разнообразные взаимосвязи, складывающиеся между филогенетически далекими видами в природных сообществах - биогеоценозах.
По определению В. А. Ковды, биогеоценоз (или экосистема) представляет собой исторически сложившуюся совокупность растений, животных, почвы, рельефа, климата и вод в данном географическом районе. Нас интересуют здесь живые компоненты биогеоценоза, т.е. входящие в его состав виды организмов, сообщество которых называют биоценозом.
(Экосистема - термин, часто используемый как синоним биогеоценоза, имеет более широкое значение, поскольку применяется также для обозначения подсистем последнего и для искусственных комплексов организмов, например, аквариум с его обитателями).
Элементарными функциональными единицами биоценоза являются популяции разных видов организмов, группирующиеся в так называемые консорции - совокупности популяций, объединяемых трофическими (пищевыми) связями с определенными видами автотрофных растений. Органические вещества, синтезируемые растениями, и аккумулированная в этих веществах энергия передаются в консорциях по трофической цепи, включающей растения (продуценты), растительноядных животных (первичные потребители), хищников (вторичные потребители) и различных сапрофагов (третичные потребители -- трупоеды, копрофаги и т.п.). Конкретная величина потока энергии и органических веществ, передающегося по трофической цепи, определяет численность популяций каждого вида организмов, входящего в состав данной консорции. Биомасса первичных, вторичных и третичных потребителей вместе составляет около 1% биомассы продуцентов (растений).
Под влиянием естественного отбора биоценозы приобретают характер устойчивых саморегулирующихся систем с обратной связью, регуляция которых - ценотический гомеостаз - осуществляется посредством вчутрипопуляционных механизмов (отдельно для популяций каждого вида) и на уровне целого биоценоза путем так называемой канализации трофических цепей, под которой понимается повышение их устойчивости за счет развития резервных "обходных путей" для передачи потоков вещества и энергии при нарушении каких-либо звеньев основной цепи. Наличие механизмов саморегуляции придает биоценозу определенную целостность. Изменения популяции одного вида в той или иной степени сказываются на популяциях других видов, входящих в тот же биоценоз. Следовательно, эволюционные изменения каждого вида затрагивают и другие виды, связанные с первым в данном биоценозе и, благодаря связям разных биоценозов друг с другом, могут в какой-то степени отражаться и на популяциях некоторых видов, входящих в другие биоценозы. Кроме того, разные популяции одного вида обычно входят в состав разных биоценозов. Эволюция видов, входящих в один биоценоз, является согласованной (или "когерентной", по В. А. Красилову). Эволюция видов организмов тесно связана с эволюцией соответствующих биоценозов как целостных биологических макросистем. При этом биоценоз может быть более устойчивой системой, чем входящие в его состав отдельные виды. Механизмы ценотического гомеостаза позволяют биоценозам сохранять стабильность, несмотря на выпадение из их состава отдельных видов или их замещение новыми.
Положение и функциональная роль данного вида в биогеоценозе характеризуются в экологии понятием "экологическая ниша". Экологическая ниша определяется, с одной стороны, составом и й данного сообщества (в этом аспекте экологическая ниша представляет собой совокупность различных ресурсов, обеспечивающих возможность существования определенного вида в данном биогеоценозе), с другой - особенностями биологии данного вида. В соответствии с этим каждый вид организмов занимает в биогеоценозе определенную экологическую нишу, но не обязательно все ниши в ценозе заняты в данный момент его исторического развития.
Судьба новых видов, внедряющихся в уже сложившееся сообщество, и их воздействие на виды, входившие в его состав прежде, определяются наличием или отсутствием в биоценозе свободных (или хотя бы используемых не полностью) экологических ниш. Нередко интродукция нового вида приводит к угнетению и даже вымиранию тех или иных местных видов. Например, хорошо известно угнетающее воздействие на многие местные виды в биоценозах островов Тихого океана животных, завезенных на эти острова человеком, - черных и серых крыс, одичавших свиней, коз, собак и кошек. С другой стороны, известны случаи, когда новые виды широко распространяются и приобретают значительную численность в новых местообитаниях, не вызывая существенных нарушений в соответствующих биоценозах. Эффектный пример такого рода представляет широкое расселение египетской цапли (Bubulcus ibis) в биоценозах Южной, а затем Северной Америки. Предполагают, что "популяция основателей" египетской цапли была занесена из Старого Света в Америку в 80-е гг. XIX в. ураганом. Египетская цапля не конкурирует ни с какими американскими видами цаплевых, так как кормится главным образом насекомыми и другими беспозвоночными, собираемыми среди травы в наземных местообитаниях, тогда как большинство других видов цапель добывает разнообразную животную пищу (в основном позвоночных) на мелководье. До появления египетской цапли в Америке соответствующая экологическая ниша в местных биоценозах оставалась незанятой.
Проблемы, связанные с эволюционной ролью биоценозов, по сути дела, лишь намечены в современной науке. Раздел экологии, изучающий биоценозы и их эволюцию (биоценология), переживает бурное развитие. Установлены, в частности, некоторые общие тенденции эволюции биоценозов и определенные циклические изменения, происходящие в процессе этой эволюции (сукцессия биоценозов). "Молодые" биоценозы характеризуются относительно малым количеством составляющих их видов, биопродукция органических веществ в них намного превышает деструкцию, т.е. разрушение органических веществ. В молодых биоценозах преобладают силы движущего отбора, обусловливающие относительно быстрые эволюционные изменения. Постепенно
число видов в биоценозе возрастает, усложняются взаимосвязи различных популяций и увеличивается количество трофических цепей. Соответственно возрастают возможности саморегуляции биоценоза. В зрелых биоценозах, находящихся в состоянии климакса, биопродукция равна деструкции органического вещества, т.е. общая продукция фотосинтеза равна суммарному дыханию всей экосистемы. (Климакс - устойчивое состояние биоценоза, являющееся завершающим этапом сукцессии). В таком зрелом, хорошо сбалансированном биоценозе существует устойчивое равновесие между популяциями различных видов, "притертых" естественным отбором друг к другу. Это благоприятствует действию стабилизирующего отбора, который оказывает тормозящее воздействие на эволюцию.
Вместе с тем Л. Ван Вален и ряд других ученых полагают, что биотические компоненты природных экосистем продолжают эволюционировать и при остающихся стабильными абиотических условиях, поскольку достигнутая степень приспособленности организмов обычно относительна и для сохранения любым данным видом его положения в сообществе необходима непрерывная адаптивная эволюция, в процессе которой и поддерживается колеблющееся равновесие между популяциями разных видов. Эта концепция получила название "гипотезы Черной (или Красной, в англоязычной литературе) королевы" - по соответствующему персонажу сказки Л.Кэррола "Алиса в Зазеркалье", утверждавшему, что "нужно бежать изо всех сил, чтобы оставаться на том же месте".
Внутри биоценозов имеются пары и группы видов, связанных друг с другом более тесно, чем с остальными членами сообщества. Прежде всего это виды организмов, принадлежащие к одной консорции. Однако взаимоотношения разных видов очень разнообразны и отнюдь не сводятся к трофическим связям (например, использование организмами одного вида нор, гнезд или тела особей другого вида как среды обитания; мимикрия, "гнездовой паразитизм", подобный таковому у кукушек, и т.п.). Эволюционные взаимодействия видов организмов, не обменивающихся друг с Другом генетической информацией, но тесно взаимосвязанных биологически, называют коэволюцией. В сущности, эволюция любого вида организмов протекает как коэволюция с теми или другими видами - партнерами по биоценозу (растения и Растительноядные животные, хищники и их жертвы, паразиты и их хозяева, различные формы симбиоза-мутуализма и др.). Многие черты конкретных филогенетических преобразований различных видов организмов можно понять, только учитывая их коэволюционные взаимосвязи. Один из наиболее известных и простых примеров этого - развитие цветка у опыляемых насекомыми покрытосеменных растений.
Поскольку развитие взаимных приспособлений благоприятно для обоих взаимодействующих видов, отбор способствует коэво-люционным изменениям. При кажущейся антагонистичности членов таких коэволюционных пар, как хищник - жертва или паразит - хозяин, коэволюция всегда обусловлена совместимостью партнеров в биоценозе и ведет к развитию таких взаимоотношений между ними, при которых виды-партнеры становятся взаимно необходимыми. Многочисленные работы экологов убедительно показали, что хищники и паразиты, регулирующие численность популяций своих жертв и выбраковывающие среди них в первую очередь больных, ослабленных, генетически неполноценных особей, являются необходимым фактором, обеспечивающим оптимальное состояние популяций видов-жертв.
Коэволюционные межвидовые взаимодействия могут продолжаться длительное время, приводя к глубоким изменениям взаимодействующих видов.
Разрушение биоценоза благодаря каким-либо внешним воздействиям (изменения условий внешней среды, превосходящие возможности саморегуляции сообщества) приводит к распаду сложившихся систем межвидовых взаимодействий и открывает простор для действия интенсивного движущего отбора, который может принять характер катастрофического. При этом утрачивается регулирующее воздействие биоценоза на эволюцию входящих в его состав видов, которая становится несогласованной ("некогерентной", по В. А. Красилову). Темпы видообразования в отдельных филетических линиях могут при этом значительно возрастать.
Некоторые исследователи (В.В.Жерихин, А.С.Раутиан и др.) полагают, что биоценотические кризисы могут возникать и без существенных изменений абиотических факторов внешней среды, развиваясь на основе специфических взаимодействий входящих в сообщество видов организмов, которые эволюционируют и в относительно стабильных условиях в соответствии с "принципом Черной королевы", по Ван Валену. Кризис сообщества может возникнуть в результате прогрессирующей и далеко зашедшей (при длительно остающихся стабильными внешних условиях) специализации большинства видов данного биотического комплекса, снижающей устойчивость сообщества. Относительно кратковременные неблагоприятные изменения внешних условий, которые могут выдержать сбалансированные биоценозы, стимулируют начало кризисных процессов в сообществах, состоящих в основном из специализированных видов при дефиците видов-универсалов, которые могли бы занять в трофических цепях места, освободившиеся при вымирании специализированных форм.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 1396; Нарушение авторских прав?; Мы поможем в написании вашей работы!