КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Проводники болевой чувствительности
|
|
|
|
Афферентные волокна групп Аδ и С — являются периферическими отростками первичных чувствительных нейронов, тела которых расположены в спинальных ганглиях. Центральные аксоны этих нейронов входят в спинной мозг в составе дорсальных корешков.
Однако точка контакта Аδ и С волокон со вторым нейроном различна.
Афферентные волокна групп Аδ контактирует с наиболее периферическими нейронами дорсального рога — нейронами первой пластинки. Нейроны первой пластинки отвечают преимущественно на ноцицептивные стимулы, поэтому их называют специфическими ноцицептивными нейронами.
Афферентные волокна групп С проникают в глубь серого вещества и оканчиваются на нейронах основания дорсального рога — главным образом на нейронах второй и третьей пластинок, которые отвечают как на ноцицептивные, так и на неноцицептивные — механические и термические раздражения.
Есть еще одна особенность прежде всего С волокон - войдя в мозг, каждое волокно делится на две ветви — восходящую и нисходящую. Каждая из этих ветвей соответственно поднимается или опускается на один-три сегмента - тракт Лиссауэра, после чего образуются синаптические контакты с нейронами дорсального рога. Это приводит к тому, что боль от С волокон плохо подается чёткой локализации.
Медиаторы ноцицептивных нейронов спинного мозга
Передача ноцицептивной информации с первичных афферентных волокон на вторые нейроны дорсального рога в спинном мозге осуществляется при участии различных нейромедиаторов — возбуждающих аминокислот (глутамат, аспартат) и нейропептид (вещество Р).
Для глутамата на постсинаптической мембране 2 нейрона есть как AMPA-рецепторы, так и NMDA-рецепторы.
С помощью этих медиаторов возбуждается второй нейрон, который будет формировать спиноталамический восходящий путь – главный путь проведения болевой чувствительности.
Пути проведения болевой чувствительности.
Аксоны второго нейрона в дорсальных рогах переходят на противоположную сторону в составе передней комиссуры, расположенной кпереди от центрального канала. Затем они направляются в головной мозг, формируя спиноталамический тракт, занимающий переднебоковые отделы белого вещества спинного мозга.
Эволюционно и функционально спиноталамический тракт делят на 2 части:
новый — неоспиноталамический, расположенный более латерально,
старый — палеоспиноталамический, расположенный медиально.
Неоспиноталамический тракт формируется в основном за счёт нейронов, которые получили информацию от Аδ афферентов, а палеоспиноталамический тракт – от С волокон.
Волокна неоспиноталамического тракта, перейдя на противоположную сторону, оканчиваются преимущественно в вентробазальных ядрах таламуса, нейроны которых проецируются в соматосенсорную кору больших полушарий.
Активация неоспиноталамического тракта — латеральной системы боли — ответственна как за само ощущение, так и за качественную оценку (характеристики боли) и локализацию боли.
Восходящие волокна палеоспиноталамического тракта оканчиваются первоначально на нейронах ретикулярной формации ствола мозга, которые посылают аксоны к интраламинарным ядрам таламуса. Нейроны интраламинарных ядер таламуса проецируются в лимбическую и лобную кору. Палеоспиноталамический тракт является, поэтому, на самом деле спиноретикулоталамическим трактом. Активация палеоспиноталамического тракта (медиальной системы боли) ответственна за такие компоненты боли как аффективный (эмоциональный) – лимбическая система и мотивационный – лобная кора. Кроме того, в норме связь с ретикулярной формацией и интраламинарными ядрами таламуса, которые участвуют в формировании «системы пробуждения» после сна объясняет, почему человек испытывающий боль не может уснуть.
Небольшая часть восходящих аксонов ноцицептивных нейронов спинного мозга, примыкающих к спино-таламическому тракту, оканчивается в верхних и нижних буграх четверохолмия, формируя спинотектальный тракт. Нейроны этого тракта участвуют в модуляции боли и в интеграции визуальной и соматической сенсорной информации.
Итак, нео спиноталамический тракт переключается в вентробазальных ядрах таламуса и нейроны этих ядер оканчиваются в соматосенсорной коре больших полушарий, в результате формируется ощущение боли, которому мы можем дать качественную характеристику и локализовать.
Палео спиноталамический тракт широко оканчивается в стволовых отделах:
- В ретикулярной формации, из нее отростки направляются в интерламинарные ядра таламуса, а от туда в лимбическую кору – возникает негативное эмоциональное состояние и в лобную кору – возникает мотивация избавить себя от боли (например, сходить к врачу). Вдобавок нейроны ретикулярной формации и интерламинарных ядер участвуют в пробуждении, а потому с болью легко проснуться и тяжело уснуть.
- В буграх четверохолмия, благодаря чему формируется чётко образ источника боли и создается условный рефлекс избегания этого источника.
В самой коре чёткого представительства боли нет.
Мелкий шрифт можно не читать – это дополнительный материал для тех, кто хочет стать челюстнолицевым хирургом.
Ноцицептивные волокна групп Аδ и С, иннервирующие органы передней части головы, входят в состав тройничного, лицевого, языкоглоточного и блуждающего нервов. Все они являются периферическими отростками нейронов, тела которых расположены в чувствительных ганглиях соответствующих черепно-мозговых нервов. Центральные отростки этих нейронов входят в ствол мозга и направляются книзу, образуя так называемый спинальный тракт тройничного нерва, занимающий латеральные участки нижнего отдела моста, продолговатого мозга и двух верхних шейных сегментов спинного мозга. Волокна этого тракта оканчиваются на нейронах спинального ядра тройничного нерва, которое расположено рядом, медиальнее самого тракта. Аксоны ноцицептивных нейронов спинального ядра тройничного нерва переходят на противоположную сторону через ретикулярную формацию ствола мозга, поднимаются и оканчиваются в задних и задневентральных отделах таламуса, образуя передний тригеминоталамический тракт. Таламические нейроны, возбуждаемые импульсами в тригеминоталамическом тракте, проецируются в соматосенсорную кору. Тригеминоталамический тракт, который относится к латеральной системе боли, обеспечивает появление острой, хорошо локализованной боли. Диффузная, плохо локализованная боль, ощущаемая в передних отделах головы, обусловлена активацией тригеминоретикулоталамического пути, который является частью медиальной системы боли. Аксоны вторичных нейронов этого пути оканчиваются на нейронах ретикулярной формации ствола мозга. Нейроны ретикулярной формации посылают аксоны к интраламинарным ядрам таламуса, нейроны которых имеют диффузные проекции в коре мозга.
Модуляция боли
Среди механизмов модулирующих боль наиболее известным является воротный механизм.
В этом ряду стоит «воротная теория» боли (Мелзак, Уолл), придающая большое значение в формировании болевых ощущений нейронам желатинозной субстанции спинного мозга.
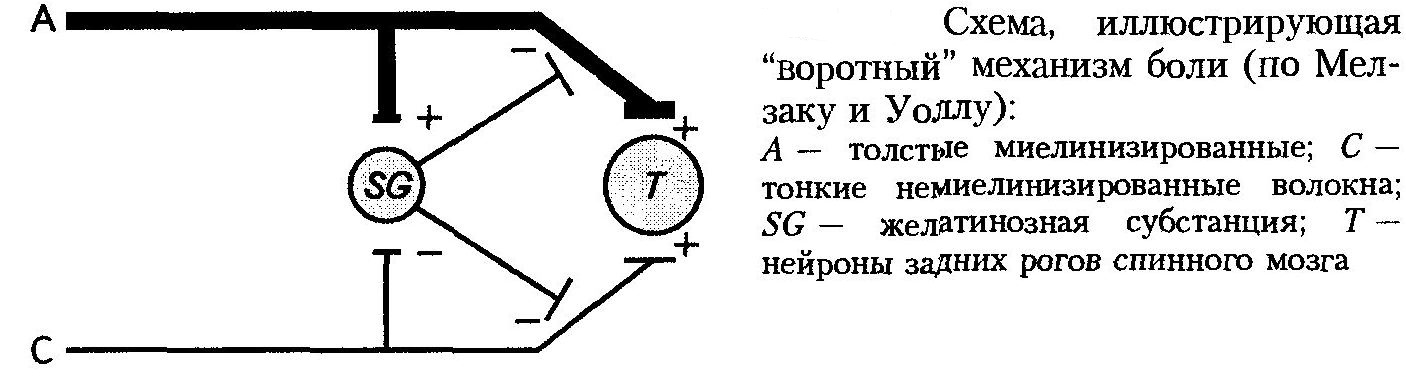
Нейроны желатинозной субстанции осуществляют пресинаптическое торможение, блокируя прохождение импульсов в нейроны задних рогов спинного мозга по толстым и тонким нервным волокнам. Если нейроны желатинозной субстанции возбуждаются, происходит пресинаптическое торможение — «ворота» закрыты. Если нейроны желатинозной субстанции сами заторможены, то пресинаптическое торможение снимается — «ворота» открыты.
Интенсивная стимуляция толстых миелинизированных нервных волокон вызывает возбуждение нейронов желатинозной субстанции — «ворота» закрываются, и проведение импульсов в спинной мозг уменьшается.
При интенсивном возбуждении тонких немиелинизированных волокон, проводящих боль, происходит торможение нейронов желатинозной субстанции, снимается пресинаптическое торможение и облегчается поступление импульсов в задние рога спинного мозга.
С нейронами желатинозной субстанции осуществляющих торможение имеют связи и другие отростки, например механорецепторов – поэтому механическое воздействие, физиопроцедуры уменьшают боль. Кроме того, нейроны желатинозной субстанции имеют нисходящие связи от центральных антиноцицептивных структур.
|
|
|
|
|
Дата добавления: 2015-03-29; Просмотров: 1996; Нарушение авторских прав?; Мы поможем в написании вашей работы!