КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Н - глу Арг энкефалины лей тре тре лей глу ом 2 страница
|
|
|
|
- полимодальные рецепторы, связанные преимущественно с С-волок-нами возбуждаются механическими, температурными и химическими раздражителями.
И всё же болевой рецептор, по своей сути, должен являться хеморецептором, так как любое повреждение тканей приводит к изменению химических компонентов окружения и вызывает появление алгогенных веществ, которые могут быть разделены на три группы: тканевые - серотонин, гистамин, ацетилхолин, некоторые простагландины, ионы калия и водорода; плазменные - брадикинин, каллидин; выделяющиеся в нервных окончаниях, например, субстанция Р (она вероятно действует как синаптический передатчик в задних рогax спинного мозга). Именно эти химические соединения, вызывают деполяризацию мембран, формируют нервный импульс.
Хеморепторыимеются в коже и слизистых оболочхах, но особенно много их вo внутренних органах, где они располагаются в стенках мелких артерий. От большинства из них импульсы про-водятся по С-волокнам иформируют протопатическую боль. Ведущим механизмом возбуждения хемоноцицепторов является гипоксия. Следовательно любой типовой патологический процесс, создающий в тканях дефицит кислорода и нарушающий процессы биологического окисления, (травмы, воспаление, ишемизация и т.д.), будет сопровождаться болью.
Такие медиаторы боли, как гистамин, простагландины (Е2 и F2а), серотонин, кинины (брадикинин), тахикинины (субстанция Р), выработка которых каскадно нарастает при травме, действуют двояко. Предполагается, что с одной стороны они облегчают возбуждение ноцицепторов, а с другой сами частично обладают возбуждающим эффектом. В конечном итоге, их появление в тканях в дополнительных количествах способствует усилению боли. Вероятно, есть какая-то дифференциация в действии медиаторов боли, поскольку их выработка зависит от характера повреждающего агента:
- механический стимул способствует выделению серотонина и брадикинина;
- химическое воздействие, вызывающее боль, опосредуется простагландином Е2;
- поверхностный термальный стимул вызывает выделение брадикинина, а простагландины в этом механизме вообще не участвуют.
Вероятно, эта дифференциация имеет место ив последующих нейроналных переключениях.
Основными свойствами ноцицепторов являются возбудимость, сенсибилизация и отсутсвие адаптации. Они относятся к высокопороговым рецепторам и возбуждаются только при действии факторов, cпocoбных вызвать разрушение тканей, однако, следует подчеркнуть,что уровень этого порога может меняться. Особенно высок порог болевой чувствительности во внутренних органах, которые не реагируют на укол или разрез, а только на растяжение.
2.3.Проводники болевой чувствительности.
Сформировшиеся пpи возбуждении ноцицепторов импульсы, проводятся по афферентным во-локнам периферической нервной системы. В их проведении участвуют два типа нервных волокон, которые, вероятно, являются собственно ноцицептивными:
- А-дельта волокна - тонкие миелинизированные структуры, проводящие боль и импульсы теплового воздействия со скоростью 12-30м/сек. Они являются гистаминергическими и accoциир-уются с первичной, острой, чётко локализованной фазной болью. Они проводят большой oбъём информации, чем С-волокна.
- С-волокна - немиелинизированные соматические и постганглионарные симпатические во-локна. Они обладают средней скоростью проведения 0,25-2 м/сек. С-волокна принимают участие в передаче ноцицептивных сигналов от висцеральных органов, приводя вторичную, плохо локализо-ванную, ноющую, жгучую и длительную боль.
Среди других периферических волокон, принимающих участие в формировании ноцицептив-ных oщущений можно выделить А - β волокна - толстые миелинизированные образования со скоростью проведения нервнцых импульсов 30-70 м/сек. Они являются ацетилхолинергическими и принимают участие в механизмах "системы воротного контроля".
2.4. Роль структур спинного мозга в ноцицепции.
По нервным проводникам ноцицептивная информация достигает прежде всего пластин серого вещества задних рогов спинного мозга, именуемых слоями Рекстеда. Здесь располагается первая релейная станция рефлекторной болевой дуги. Переключение болевой импульсации происходит при участии химических медиаторов, которые могут оказывать противоположные влияния на про-хождение болевого потока как стимулирующе, так и тормозящие:
- типичным представителем первой группы является субстанция P, высвобождение которой приводит к резкому увеличению прохождения потока болевой информации в мозг. Этот процecc неразрывно связан с образованием, освобождением и выделением других медиаторов боли, усиливавших алгогенный эффект: гистамина, ацетилхолина, серотонина, кининов, простагландинов. При этом острая локализованная боль постепенно заменяется наразлитую или протопатическую.
- медиаторы другой группы, такие как глицин, серотонин, норадреналин, дофамин, энкефалины, оказывают тормозящее влияние на ноцицептивные структуры и замедляют прохождение болевых сигналов.
С у б с т а н ц и я P - важнейший медиатор боли. Помимо задних рогов спинного мозга, она обнаружена также в стволе, преоптической области гипоталамуса и, в небольших количествах, в коре голoвного мозга, то есть на всех участках релейной передачи болевых импульсов. Субстан-ция P накапливается в синапсах и освобождается при деполяризации. Она является полипепти-дом, который по структуре может быть отнесён к катехоламинам. Предполагается, что субстанция P обезпечивает преимущественное проведение болевых импульсов при механических воздействиях. Введение пептида к а п с а и ц и н а увеличивает освобождение субстанции, истощая её запасы, что повышает порог болевой чувствительности и формирует аналгезию. Участие субстанции Р в передаче болевой информации блокируется эндогенными опиоидами, морфином и морфиноподобными веществами, что, вероятно, связано с воздействием на пресинаптическую мембрану.
Нейроны задних рогов помимо вышеперечисленных медиаторов возбуждаются под влиянием холецистокинина и нейротензина, а тормозятся - γ-аминомаслянной кислотой, соматостатином.
2.5. Роль проводящих путей и других структур центральной нервной системы в ноцицепции.
Ноцецептивная информация из задних рогов спинного мозга поступает в вышерасположенные отделы головного мозга по восходящим ноцицептивным проводящим путям: спино-таламическо-
му, спино-ретикулярному и спино-мезэнцефалическому трактам. Они проходят и сосредоточены в передне-боковом квадранте спинного мозга. Их перерезка (хордотомия) приводит к аналгезии, хотя в дальнейшем болевое чувство может восстановиться. Это говорит о том, что в передаче боли, вероятно, принимают участие и другие проводящие пути.
Прохождение болевых импульсов по разным проводящим путям предполагает завершение их вразличных структурных образованиях головного мозга, что обуславливает формирование отли-чающихся друг от друга проявлений боли и сопутствующих эффектов. Их можно обозначать, как компоненты болевой реакции:
- первый компонент - двигательная защитная реакция, возникает обычно даже до появленя основного ощущения боли. Она формируется в результате распространения болевого импульса в спинном мозге на мотонейроны.
- специфический сенсорный путь по лемнисковым nутям и медиальной петле проводит им-пульс в заднюю группу ядер гипоталамуса. Здесь происходит второе переключение, после чего импульсы поступают на нейроны соматосенсорной зоны коры в поле S1, реакция которых и проявляется во второй системной реакции - болевом ощущении. Здесь формируется ощущение эпикритической боли, а поскольку oнa возникает, как пpaвило, неожиданно, то поведенческая реакция проявляется во вздрагивании и настораживании;
- неспецифические экстралемнисковые пути не имеют чётко определенных морфологических границ. Они описываются как спино-ретикулярные пути, которые заканчиваются в различных нейрональных структурах,в частности, при попадании ноцицептивного потока нанейроны ретикулярной формации ствола мозга, происходит генерализация реакций с возбуждением нижe и выше расположенных структур. Тут формируется третий системный компонент боли – п р о б у ж - д е н и е - б о д р с т в о в а н и е (arousal-реакиия), которая может быть снята наркозом.
Помимо этого, импульсация, проходящая по этим же путям, может активировать нейроны ги-поталамуса - высшего вегетативного центра. Это обусловливает проявления четвёртого по счёту компонента системной болевой реакции - в е г е т а т и в н о г о: повышение артериального давления, увеличение частоты дыхания и числа сердечных сокращений, увеличение выделения гормонов, в том числе кортикотропина, выброс гормонов тревоги - адреналина, норадреналина. Всё это в совокупности в конечном итоге, влияя на скорость метаболических процессов, приводит к перестройке обмена веществ.
Сочетанная активация ретикулярной формация и гипоталамуса, отвечающих за эмоции, приводит к появлению пятого компонента системной болевой реакции – о т р и ц а т е л ь н ы х э м о ц и й. Следует отметить, что вегетативный и эмоциональный компоненты системных реак-ций могут быть заблокированы введением транквилизаторов, но само ощущение боли при этом сохраняется.
С активацией фронтальной и, по-видимому, теменной областей коры больших полушарий связано формирование шестого компонента системной болевой реакции - мотивационого избавления от боли. Замечено, что удаление лобных долей коры у человека вызывает безразличное отношение к болевому ощущению, хотя сама боль при этом может даже усилиться. Поражение теменных долей приводит к"болевой асимволии", характеризующейся отсутствием психических реакций на боль.
Таким образом очевидно, что всё многообразие проявлений при болевом синдроме формируется при участии самых различных структурных образований головного мозга. Функциональная роль каждого из них была выяснена как в эксперименте, так и в ходе клинических наблюдений за больными с поражением отдельных участков мозга. Было, например, установлено, что нейроны гиганто-клеточного ядра влияют на прохождение болевой импульсации: при его стимуляции боль усиливается, а при повреждении нейронов этого ядра прохождение ноцицептивных импульсов тормозится. Поражение ядер гипоталамуса сопровождается появлением крайне мучительных жгучих болей в области лица, дистальных отделах рук и ног, возникающих с наступлением сна. Нивелирование этих болей седуксеном свидетельствует о том, что здесь имеет место нарушение фильт-
Основные проявления Ноцицептивная Антиноцицептивная
боли система система
 Болевой раздражитель I Сегмантарный уровень
Болевой раздражитель I Сегмантарный уровень
контроля боли
Повреждение. Раздражение II Нисходящая аналгетичес-
 болевых рецепторов кая система
болевых рецепторов кая система
 |
 Образование субстанции P 1. Кора - задние рога
Образование субстанции P 1. Кора - задние рога

 Повышение чувствитель- и других алгогенов 2. ЦСОВ - ядра шва
Повышение чувствитель- и других алгогенов 2. ЦСОВ - ядра шва
 ности болевых рецепторов 3. ЦСОВ - голубое пятно,
ности болевых рецепторов 3. ЦСОВ - голубое пятно,
 Возбуждение А-дельта 4. Гипоталамический уро-
Возбуждение А-дельта 4. Гипоталамический уро-
и С-волокон вень контроля

 5. Корковый уровень конт-
5. Корковый уровень конт-
роля


 Двигательная реакция Возбуждение желатинозной 6. Опиатный механизм
Двигательная реакция Возбуждение желатинозной 6. Опиатный механизм

 сегментарного уровня субстанции и клеток 7. Норадренергический
сегментарного уровня субстанции и клеток 7. Норадренергический
I – VI пластин заднего рога механизм
 8. Холинергический
8. Холинергический
механизм
 9. ГАМК-ергический
9. ГАМК-ергический
Проведение возбуждения механизм
по спино-таламическому 10. Серотонинергический
тракту и короткоаксонным механизмы
 путям спинного мозга
путям спинного мозга
 | |||
 | |||
гигантоклеточное ядро

голубое ядро
Активация САС Болевая информация
 и ГГНС от области головы ядра шва
и ГГНС от области головы ядра шва

ЦСОВ
 | |||
 | |||
Formatio reticularis


гипоталамус

 Вегетативные реакции
Вегетативные реакции
 задние ядра
задние ядра
 Эмоциональные реакции медиальные ядра
Эмоциональные реакции медиальные ядра
 | |||||
 |  | ||||
таламус
Двигательные реакции Соматоненсорная Лимбические Диффузная активация
 супраспинального уровня область коры образования коры
супраспинального уровня область коры образования коры
Поведенческие Острая локализованная Острая нелокализованная

 реакции боль боль
реакции боль боль
Память Острая боль
рации импульсов не только в гипоталамусе, но и в ретикулярной формации. Роль лимбических
структур в формировании эмоционально-поведенческих компонентов боли: психического возбуждения, ярости, агрессии, страха, гнева подтверждается тем, что после перерезки нервных путей, соединяющих таламус с лимбическими образованиями (лейкотомии), развивается полное безразличие к боли, хотя сама боль сохраняется.
3.1. Структурные основы и функциональные особенности антиноцицептивной системы.
Несмотря на обилие теоретических концепций, по мере накопления клинических и научных наблюдений, всё большее числo проявлений боли не вписывались в классические представления о её механизмах. Например, не находила объяснения врожденная аналгезия при полностью сохраненных проводящих путях и центрах болевой чувствительности; не понятен был механизм возникновения, так называемого синдрома "депрессия в маске", при котором люди ощущали боль в сердце, зубах или других органах, при полном объективном благополучии в них. Удовлетворительные объяснение эти явления получили только после того, как были сформулированы основные положения антиноцицептивной концепции, возникновение и формирование которой было продиктовано и подготовлено объективным ходом развития событий.
Впервые в 1969 году американский физиолог Reynolds при возбуждающих воздействиях на некоторые точкицентрального серого вещества зарегистрировал у бодрствующих животных феномен аналгезии - отсутствие реакции на болевые раздражения лап, хвоста, брюшины и других частей тела, при этом не отмечались изменения других сенсорных реакций. Всё это свидетельствовало в пользу чистой анальгезии, а не анестезии. В последующем аналогичные результаты были получены и у людей. Дальнейшие исследования выявили целую сеть точек в гипоталамусе, тала-мусе, чёрной субстанции, гигантоклеточном ядре и других областях, стимуляция которых вызывала аналгезию. Выяснялось, что paздpaжeниe этих структур вызывает торможение пре- и постсинаптической ноцицептивной импульсации на релейных станциях, начиная с нейронов эадних рогов спинного мозга. Эти данные послужили основой для становления и последующего развития учения об эндогенной противоболевой системе. Наиболее заманчивым направлением при изучение особенностей функционирования этой системы предполагалось выяснение нейрохимических механизмов торможения боли, так как в этом случае медицина получила бы на вооружение аналгезирующие вещества физиологического происхождения.
В настоящее время накопленный фактический материал позволяет суверенностью утверждать, что система боли, как и другие механизмы поддержания гомеостаза, представлена двумя противоположно функционирующими структурными образованиями: алгогенными и антиноцицептивными. Причём последней отводилась роль регулятора уровня восприятия болевого ощущения. В соответствии с этой концепцией интенсивность боли предопределяется не только масштабами повреждения, но и функциональным состоянием различных отделов антиноцицептивной системы. При недостаточности отдельных её звеньев понижается порог болевого восприятия, что приводит к нарастанию интенсивности боли. Предполагается, что отсутствие боли на неопасное для организма повреждение обусловлено тем, что постоянно функционирующая антиноцицептивная система препятствует прохождению сформированных в этих условиях импульсов в вышестоящие отделы центральной нервной системы, чем и предупреждается возникновение боли. В процессе эволюции шло постепенное надстраивание различых уровней антиноцицептивной систомы, вплоть до корковых образований. Ноцицептивный и амтиноцицептный компоненты болевой системы имеют различные нейрональные механизмы, но при этом они тесно взаимодействуют друг сдругом, а соотношением их функциональной активности предопределяется возникновением различных вариантов болевого синдрома.
Г и п е р а л г е з и ю можно вызвать либо повышением активности ноцицептивной системы - например, введением её медиатора брадикинина; ли6о снижением активности антиноцицептивной системы - введением антисыворотки к опиоидным пептидам или блокаторов образования пред-шественников серотонина. Предполагают, что именно этот механизм лежит в основе развития мигрени. Различные варианты воздействия на компоненты антиноцицептивной системы позволяют получать в экспериментах на животных боли различной интенсивности. Например, охлаждение, отменяющее влияние нисходящих антиноцицептивных путей, вызывает не только гипералге-
зию, но и спонтанные боли. Введение в сегмент спинного мoзга иннервирующий заднюю конечность, столбнячного токсина, блокирующего все медиаторы антиноцицепции, вызывает такие боли, что крыса отгрызает конечность.
Г и п о а л г е з и ю можно выявить повышением активности антиноцицептивной системы - введением морфия или акупунктурой; следует отметить, что врожденная аналгия обусловлена генетически высоким уровнем активности антиноцицептивной системы.
Эволюционно сформировался такой уровень взаимодействия этих двух противоположных систем, который является целесообразным с позиции сохранения жизненно-важных констант организма.
На различных уровнях ЦHC антиноцицептивная система имеет свои морфологические, физиологические и биохимические структуpы и механизмы. В совокупности аитиноцицептивных механизмов последние годы сделаны попытки выделить четыре основные аналгетические системы:
- нейрональную-опиатную - энкофолинэргическую;
- нейрональную-неопиатную - норадреналин-,серотонин-,дофаминергическую;
- гормональную-опиатную - эндорфиновую;
- гормональную-неопиатную - пептиды и другие регуляторы.
Все эти системы - опиатная, моноаминергическая, холинергическая, ГАМКергиская, гормона-льная и нервная функционируют не изолированно, а тесном взаимодействии между собой и другими системами.
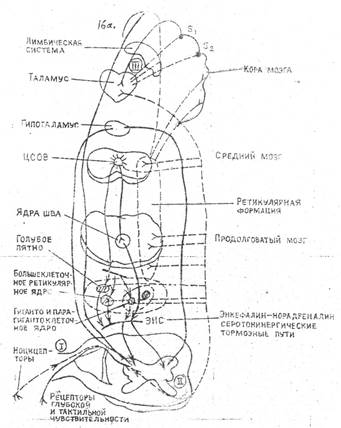
Рис-2. Ппримерная схема нисходящих и других антиноцицептивных тормозных путей.
------ болевые пути (I, II, III, нейроны болевых путей)
 нисходящие и другие пути
нисходящие и другие пути
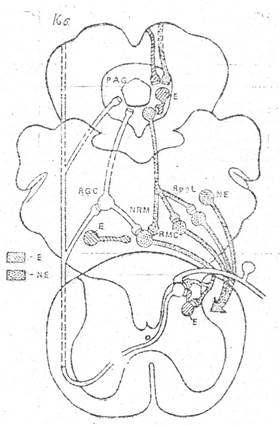
Рис- 3. Схема главных компонентов нисходящей антиноцицептивной схемы ствола мозга
(по A. Basbaum, H. Field, 1984).
PAG – околоводопроводное серое вещество, RGC – ретикулярное гигантоклеточное ядро,
Rpgl – ретикулярное парагигантоклеточное, NRM – большое ядро шва,
RMC – ретикулярное крупноклеточное ядро, E – нейронные системы, содержащие опиоидные пептиды, NE – норадренергическая и серотонинергическая системы.
Каждое из структурных образований ЦНС вносят свой вклад в формирование ноци-антиноци-цептивных oтношений:
- гипоталамусу принадлежит важная роль в синтезе и секреции энкефалинов и эндорфинов, а также в усилении образования гипофизом тропных гормонов и β-липотропина, из которого и обра-зуются эндогенные опиаты гормонального происхождения, помимо этого он участвует в актива-ции симпато-адреналовой системы.
- зрительный бугор и его ядра связаны с соматосенсорной зоной, лимбическими образования-ми, двигательной идругими областями коры. С активацией этих структур связано формирование острой боли, равно как и происходит становление антиноцицептивных механизмов.
Все эти образования функционируют в тесном взаимодействии:
- импульсация, поступающая по толстым волокнам на уровень задних рогов спинного мозга повышает синтез энкефалинов, которые на сегментарном уровне уменьшают образование субстан-ции Р и тормозят передачу болевых импульсов по спиноталамическому пути;
- на уровне ствола мозга включается нисходящая аналитическая система: гиганто-клеточное ядро - ядро шва, которые посредством серотонин-, норадреналин- и энкефалинергических механизмов оказывают нисходящее влияние на задние рога и тем самым на прохождение болевой ин-формации;
- возбуждение симпато-адреналовой система тормозит передачу болевой информации и уси-ливает образование эндогенных опиатов;
- на всехрелейных участках отмечено наличие опиатных рецепторов, с которыми соединяют-ся эдогенные опиаты;
- следует учитывать также корковые влияния на все участки переключения болевой чувстви-тельности. Нарушение антиноцицептивной функции соматосенсорной зоны в коре вызывает выраженную гиперпатию;
3.2. Опиоидные механизмы антиноцицептивной активности.
Важнейшей составной частью эндогенной обезболивающей системы являются опиоидные ме-ханизмы. Отправными посылками, на основании которых была сформулирована концепция анти-ноцицептивной активности опиоидов, послужили следующие факты и открытия:
- аналгетический эффект препаратов опия, который был известен с древних времен.
- в 1973 году в лабораториях Терениуса, Шнайдера и Перта были открыты клеточные опиат-
ные рецепторы - с которыми специфически связывались морфин и его аналоги. Эти рецепторы локализовались главным обpазом в клетках ЦНС. Особенно много их оказалось на станциях переключения: в задних рогах спинного мозга, продолговатом и среднем мозге, гипоталамусе, лимбической системе, таламусе, коре больших полушарий. Вместе с тем в дальнейшем они были обнаружены и в других тканях, в частности на Т-лимфоцитах.
- в 1975 году было установлено, что в организме человека вырабатываются собственные эндогенные вещества, которые являясь по структуре олигопептидами, обладают способностью вступать во взаимодействие с опиатными рецепторами.
Эти вещества были названы энкефалинами и эндорфинами (выделенные из мозга и подобные морфину). Постоянный синтез этих пептидов в организме поддерживает в крови, спинно-мозговой жидкости и различных структурных образованиях ЦНС опроделенный уровень их концентрации. В процессе реализации своих функций они, связываясь с опиатными рецепторами, вызывают пре- и постсинаптическое торможение афферентной ноцицептивной импульсации, помимо этого они, вероятно, тормозят выделение алгогенных веществ: субстанции Р, брадикинина, простагландинов и других медиаторов боли. Любое повышение содержания этих веществ в тканях при экзогенном введении, вследствие электрораздражения антиноцицептвных структур центрального серого ве-щества, гипоталамуса, таламуса и т.д. вызывает либо гипоалгезию, либо аналгезию.
Для того, чтобы избежать терминологической путаницы, следует помнить, что:
- морфий и другие болеутоляющие соединения, содержащиеся в маке и являющиеся по своему механизму действия аналогами морфия, получили название о п и а т о в;
- вещества, близкие по фармакологическим эффектам копиатам, были названы о п и о и д а - м и или опиатоподобными веществами.
3.2.1. Структура и функция эндогенных опиоидных пептидов.
Под эндогенными опиоидами подразумеваются олигопептиды, способные вступать во взаимодействие с опиатными рецепторами нейронов и других клеток. Они содержаться в различных структурных образованиях нервной системы, помимо этого опиоидные пептиды циркулируют в крови и спинномозговой жидкости.
Предшественкиками эндогенных опиоидов является пропептиды, среди которых особая роль принадлежит проопиомеланокортину, в структуру которого как бы встроены β-эндорфин, γ-эн-дорфин, кортикотропин (АКТГ), α-, β-, γ-меланоцитстимулирующий гормоны. Синтезируется проопиомеланокортин в гипоталамусе, передней и промежуточной долях гипофиза инекоторых других тканях, находящихся под регуляторным контролем глюкокортикоидов. В аденогипофизе проопиомеланокортин расщепляется на кортикотропин и β-липотропин. В промежуточной доле кортикотропин подвергается дальнейшему pacщеплению на α- меланоцитостимулирующий гормон и кортикотропиноподобный пептид; β-липотропин - на β-эндорфин и γ-липотропин и другие активные структуры. Образование биологически активных пептидов путем последовательного протеолиза фрагментов β-липотропина хорошо иллюстрируется следующей несколько упрощенной схемой:
| |||||||||||||||||
 | |||||||||||||||||
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 465; Нарушение авторских прав?; Мы поможем в написании вашей работы!