КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Второй экскурс в эмбриологию
|
|
|
|
Читателю, знакомому с данными о психологическом развитии младенца, понятно, что в предыдущем описании трех поворотных пунктов я ограничился сообщением о непосредственных наблюдениях над младенцами и вытекающих из них психоаналитических выводах. Из эмбриологии я заимствовал лишь понятие и термин «организатор» и «силовое поле». Я говорил тут и там о некоторых аналогиях и хочу теперь отметить некоторые особенно разительные сходства в структурах эмбриологического мышления и психоаналитической теории. Я уже говорил о принадлежащей Уоддингтону (1940) концепции развития как прогрессии от нестабильности к стабильности. Она, несомненно, соответствует нашим собственным представлениям о функции психической структуры. Кроме того, концепция Уоддингтона соответствует концепции о функции идентичности Эго, разработанной Эриксоном (1956).
Далее Уоддингтон (1940) утверждает, что сам по себе стимул или отдельная причина не является адекватным объяснением чего-либо,
но что дифференциацию создает вся комплексная система действий и взаимодействий. Он говорит о способности ткани формировать определенный орган, но лишь на протяжении конкретного периода. Это во многом соответствует концепции критических периодов в психологии, продемонстрированной на собаках Скоттом и Марстоном (1950) и — независимо от их исследования — мной на младенцах (Spitz, 1958).
Не ссылаясь на формулировки Фрейда относительно комплементарных рядов и, вероятно, даже не зная о них, Уоддингтон (1940) говорит о комплементарном влиянии тканевой компетентности и эвока-тора. Между тем он признает, что имеются психологические генные воздействия. Но гораздо более важным представляется то, что, хотя он и не использует термин «критические периоды», он постоянно отмечает, что воздействие гена или определенной генной последовательности ограничено определенным периодом, в котором соответствующие гены оказывают решающее воздействие и в котором процесс разветвляется на два разных направления развития.
Эта теоретическая формулировка иллюстрируется затем на дрозофилах и домашних курах (Waddington, 1940) и даже использовалась другим автором (Goldschmidt, 1938) для формулировки гипотезы (не получившей общего признания) о половой дифференциации.
Система путей развития эволюционирует по разветвляющимся линиям. Ее равновесие непостоянно, и с течением времени состояние системы меняется. Однако нормальный путь развития — это тот, к которому развивающаяся система стремится вернуться после того, как было нарушено равновесие. Это является еще одним способом выражения эмбриологической регуляции, которая, однако, происходит лишь на протяжении определенного периода времени. Двухмерная картина разветвления в развитии вскоре становится неадекватной для описания продолжающихся процессов, и на смену ей приходит то, что Уоддингтон (1940) назвал «эпигенетическим ландшафтом», состоящим из наклонной равнины с долинами, слияниями долин и их разветвлениями.
Эти формулировки не мои; они чуть ли не дословно взяты у вышеупомянутых эмбриологов. Тем не менее они во многом звучат так, словно идеи, часто встречающиеся в психоаналитической литературе, были выражены другим способом или в несколько иной системе координат. В этом контексте является поучительным использование Уоддингтоном различных схематических средств для наглядного объяснения зависимой дифференциации. Одним из них являлся «эпигенетический ландшафт».
Другая диаграмма Уоддингтона, которую мы считаем полезной для понимания связи между организатором, зависимой дифференциацией и направлением, которое принимает развитие, воспроизведена здесь (рис. 1) в чуть измененной форме. Каждая их последовательных ступеней развития представлена конусом, соединенным с предыдущим конусом посредством организатора.
Первый организатор находится в неустойчивом равновесии на вершине верхнего конуса; он может скатиться вниз по стороне конуса в
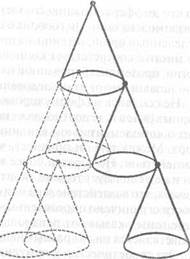
Рис. 1.
любое место на радиусе круга в 360 градусов и остановиться в одной из точек окружности, образующей основание конуса. Здесь он дает начало организатору второго порядка, второму организатору. Место, в котором он установился на окружности, необратимо определяет некоторые дальнейшие линии на пути развития.
В свою очередь второй организатор может скатиться вниз в любую точку круга в 360 градусов, который служит основанием вторичного конуса. Сам по себе вторичный конус подвижен лишь относительно той позиции, которую занимает его организатор на круговом основании первого конуса. Как только эта позиция устанавливается, положение второго конуса по отношению к первому конусу становится необратимым. Из этой необратимой точки второй организатор будет продвигаться к своей конечной позиции на основании окружности вторичного конуса; здесь сформируется третичный организатор, который в свою очередь пройдет через сходный процесс, и так далее.
Но если первый организатор скатится в другую точку круга, образующего основание первичного конуса, то сформированный там второй организатор будет индуцировать развитие в совершенно ином и новом направлении. Он будет детерминировать появление очередных организаторов, которые частично могут перекрываться, но определенно не будут совпадать с первоначальным направлением, которое было выбрано при нормальном развитии первичного организатора. Этот отличный путь представляет собой ответвление, которое необратимо будет определять в дальнейшем иную последовательность появления вторичного и третичного организаторов, как можно увидеть на диаграмме, где они изображены пунктирными линиями.
Идеи, развиваемые здесь с помощью диаграммы, во многом напоминают то, что, как я полагаю, стало для нас общепризнанным в отношении последовательных фаз либидинозного развития. Предпосылкой для нормального развития анальной стадии является более или менее ненарушенное развертывание оральной стадии. Прохождение без нарушений через анальную стадию в свою очередь создает наилучшие условия для возникновения фаллической стадии. Успешное же прохождение через эти три стадии закладывает основу для эдиповой стадии и т.д.
Если на любой из этих стадий случается серьезное нарушение, то тогда закладывается девиантная основа для установления следующей
стадии, будь то в форме точки фиксации или даже в форме более серьезных расстройств. Основываясь на впечатлениях, полученных при непосредственном наблюдении над младенцами, я бы постулировал, что это же относится к установлению организаторов психики в первый год жизни и что неадекватное установление любого из этих организаторов психики ведет к девиантному психическому развитию.
Любой внимательный читатель литературы по эмбриологии постоянно будет сталкиваться с такими аналогиями. Но, разумеется, наша концепция взаимодействия между созреванием и психологическим развитием содержит сходства, выходящие за рамки просто аналогии.
Ранее я отмечал, что вначале созреванию принадлежит большая роль, чем психологическому развитию, и что эта пропорция меняется к моменту, когда мы приходим к третьему организатору психики. Близкое сходство с процессами развития, происходящими в эпигенетическом ландшафте созревания, делает весьма вероятным, что на самой ранней стадии некоторые психологические процессы будут моделироваться по биологическим прототипам. На нынешней стадии развития нашего знания — или эмбриологического знания — вряд ли возможно провести границу между биологическими и психологическими процессами; фактически Фрейд всегда утверждал именно это.
Позвольте мне категорически подчеркнуть, что я не пытаюсь перенять концепции эмбриологической теории и просто перевести их в наши психологические термины. Во-первых, если бы даже такая попытка была желательной, наше знание о психологическом развитии все же было бы недостаточным. Уже при самом поверхностном рассмотрении становится совершенно очевидным, что область психики является намного более сложной, чем область физического созревания. Законы, управляющие созреванием, должны быть расширены и дополнены, даже если мы постулируем непрерывность континуума между сомой и психикой.
Мы можем предположить, что законы, управляющие эмбриональным развитием, будут в значительной мере применимы на протяжении нескольких месяцев и, возможно, лет после рождения. Эти законы эмбриологии будут представлять один из факторов в процессе, который мы назвали созреванием. Однако в области психологического развития любое применение этих законов будет, по существу, лишь грубой аналогией. С другой стороны, нам, конечно, известно, что природа крайне скупа на свои методы и механизмы — они представлены только что упомянутыми мною законами. Природа щедра на материал, будь этим материалом индивид, вид, род, класс или фи-люм1. Но она будет продолжать использовать тот же механизм в условиях, которые кажутся совершенно неподходящими. Поэтому было бы неудивительно обнаружить метод или механизм, используемый на эмбриональной стадии, который действует также на стадии младенчества в совсем иной среде, в среде психологического развития.
1 Единица систематики в ботанике и зоологии. ** Прим. перев.
Я бы предположил, что механизм организатора будет продолжать использоваться после рождения, по крайней мере в соматической сфере и в созревании в целом. Применяя это понятие к собственно психологическому развитию, я отдаю себе отчет, что это является аналогией и что окончательная формулировка законов психологического развития в первый год жизни — дело будущего, и она может существенно отличаться от выдвинутых мною предположений. Я выдвинул их в надежде, что они могут иметь некоторую эвристическую ценность как для нашего понимания психического развития в первые восемнадцать месяцев жизни, так и в отношении нозогенеза и нозологии.
Мы уже цитировали Оппенгеймера (1956) в поддержку использования аналогии. Однако такое использование подлежит определенным ограничениям. В своем эссе «Аналогия в науке» он также указывает, что всякий раз, когда аналогия применяется в науке, обнаруживается, что «приходится несколько расширять рамки и находить дисаналогию, которая позволяет нам сохранить то, что было верно для аналогии». Это утверждение налагает на нас обязанность указать, по крайней мере ориентировочно, дисаналогии между биологической концепцией развития и психологической.
Наиболее явная дисаналогия относится к самому понятию организатора. В биологии организатор ограничен областью бластопора, частью ткани, называемой дорсальной губой, которая устанавливает силовое поле (Needham, 1936). Ничего подобного не имеется в виду, когда мы используем термин «организатор психики». Мы мыслим в строжайшем соответствии с принципами, заложенными Фрейдом в «Я и Оно» (1923), где он представляет диаграмму психических систем. Он отрицает expressis verbis всякую возможность конкретного приложения этой диаграммы к актуальным физическим структурам. Тщетно было бы пытаться найти соматический эквивалент для организаторов психики, например в миелинизации или в аналогичных органических процессах. Психические и органические процессы взаимосвязаны; но они не являются двумя разными аспектами одного и того же феномена, они комплементарны. Грубо говоря, в тот или иной момент (мы пока еще весьма далеки от того, чтобы установить, в какой именно) миелинизация становится необходимым органическим инструментом для осуществления определенных функций, инициированных появлением организатора психики. Однако сама по себе миелинизация не является сущностью этого организатора. В другом месте (1951) я указывал на некоторые аспекты этого отношения.
Мы вынуждены допустить, что подобно тому, как понятие поля в эмбриологии, говоря словами Оппенгеймера (1956), «несколько расширяет рамки» понятия поля в физике, так и организатор психики является приложением понятия организатора в эмбриологии. Ибо, если организатор в эмбриологии — категория пространственно-временная, то организатор психики, по всей видимости, проявляется во времени, но не в пространстве.
Можно найти и необходимо исследовать и другие такие дисаналогии. Одна из них, которая тут же приходит в голову, относится к уровням организации. В эмбриологии различия между уровнями организации рассматриваются некоторыми учеными как «переход от количества к качеству» (Waddington, 1940).
«Когда элементы определенной степени сложности организуются в единицу, принадлежащую более высокому уровню организации, мы должны предположить, что когерентность более высокого уровня зависит от свойств, которыми действительно обладали изолированные элементы, но которые не могли проявиться, пока эти элементы не вступили в определенную связь друг с другом... то есть, новый уровень организации нельзя объяснить свойствами его элементарных единиц, как они ведут себя по отдельности, но можно объяснить, если мы добавим к ним определенные другие свойства, которые эти единицы проявляют, объединившись друг с другом» (Waddington, 1940).
Эта точка зрения полностью совместима с предположениями психоаналитика, касающимися функционирования психического аппарата. Но когда автор далее утверждает, что «нельзя объяснить какое-либо особое поведение, постулируя наличие нового уровня организации», то тогда мы сталкиваемся со специфической дисаналогией. Ибо в процессе психологического развития предпосылкой нового поведения действительно является образование более высокого уровня организации (или, наоборот, нарушение уже существующей организации). Остается выяснить, обусловлена ли эта дисаналогия принципиальными различиями между предметами исследования эмбриологии и психологии.
Все это — фундаментальные дисаналогии. Мы можем подумать о других — например, об экспериментах в эмбриологии, когда организатор из одной части примордия перемещают в другую часть, где он создает орган, не принадлежащий другой части, например, когда глазной бокал перемещают от переднего полюса к дорсальному эпидермису, где он превращает эпидермис в хрусталик глаза.
На нынешнем уровне развития нашего знания такие транспозиции в сфере психического функционирования невозможны. Локализация в психике не является приемлемой концепцией; согласно нашим представлениям, даже самое грубое структурное разделение психики не соответствует какой-либо физической организации в центральной нервной системе. Ее границы, границы между Ид, Эго и Супер-Эго, неустойчивы; и, как отмечала Анна Фрейд (1936), в обычных условиях граница между Эго и Супер-Эго неразличима. Она становится заметной только тогда, когда между ними возникает противоречие.
Но даже в этом случае у нас, похоже, возникают некоторые аналогии. Трансплантированный организатор, глазной бокал, создаст хрусталик из дорсального эпидермиса. Но этот глаз никогда не будет видеть, потому что у него отсутствует нервное соединение с оптическим центром мозга. С другой стороны, в силу своих имманентных законов он развивается в орган, напоминающий глаз.
Однако глаз посередине спины нарушит защитную функцию дорсального эпидермиса из-за включения более уязвимой и менее функциональной области. Все это до некоторой степени напоминает нам патологические феномены при неврозе и психозе, такие, как наличие одиночной бредовой идеи у сравнительно нормальных в остальном индивидов или ограниченного навязчивого ритуала у дееспособных в остальном людей и прочие аналогичные и внешне изолированные области дисфункции.
В эмбриологии имеется довольно много данных, подкрепляющих психоаналитические идеи. Разные авторы, среди них Беата Ранк(1949), писали о «фрагментированном Эго». Эмбриология своими экспериментами разделения дорсальной губы бластопора предоставила модель для таких феноменов. Особенно поучительным в этом отношении является эксперимент, в котором дорсальная губа бластопора разделяется на два компонента, причем один из них нежизнеспособен. Однако он будет оказывать как организующее, так и дезорганизующее влияние на компонент, который остается жизнеспособным и развивается более или менее нормальным образом. Далее, мы обнаруживаем мозаичные потенциальности у различных компонентов оплодотворенного яйца, которые выражаются в беспорядочном нагромождении квази-органов, не достигающих функциональной целостности; и мы вспоминаем о совершенно несвязной личности у некоторых психотиков.
Все это, возможно, искусственные аналогии, и нам лучше не придавать им значения. Однако нам все-таки кажется, что более широкое понятие зависимой дифференциации, в которой организаторы возрастающих порядков инициируют цепи последовательной индукции в презумптивной ткани, выходит за рамки простой аналогии. Нам также кажется, что концепция гармонической эквипотенциальности, которая гласит, что изолированным частям присуща тенденция реорганизовывать свой материал в ту же упорядоченную структуру, какой первоначально обладало целое, может быть применена и к психологическому развитию.
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 441; Нарушение авторских прав?; Мы поможем в написании вашей работы!